Comparative Metabolic Modeling of Synthetic Microbial Communities: From Ecological Theory to Biomedical Applications
This article explores the transformative role of comparative metabolic modeling in the rational design and functional optimization of Synthetic Microbial Communities (SynComs) for biomedical and biotechnological applications.
Comparative Metabolic Modeling of Synthetic Microbial Communities: From Ecological Theory to Biomedical Applications
Abstract
This article explores the transformative role of comparative metabolic modeling in the rational design and functional optimization of Synthetic Microbial Communities (SynComs) for biomedical and biotechnological applications. We provide a comprehensive analysis of the foundational ecological principles governing microbial interactions, detailing advanced methodological frameworks that integrate genome-scale metabolic models (GEMs), proteogenomics, and machine learning. The content systematically addresses critical challenges in model reliability and community stability, while evaluating validation strategies and comparative performance of different reconstruction tools. Aimed at researchers, scientists, and drug development professionals, this review synthesizes a pathway towards predictive microbiome engineering, highlighting its potential to revolutionize therapeutic development and personalized medicine.
Decoding Microbial Ecosystems: The Principles of Community Assembly and Interaction
Defining Synthetic Microbial Communities (SynComs) and Their Biomedical Promise
Synthetic Microbial Communities (SynComs) are consortia of microorganisms that are artificially combined to confer specific, beneficial functions collectively [1]. They represent a shift from single-strain microbial inoculants to a systems-focused approach, leveraging multi-microbe and host interactions that exhibit emergent properties not present in single-isolate approaches [1]. The core principle behind SynComs is to reduce the overwhelming complexity of natural microbial communities while preserving essential ecological interactions, thereby creating a more tractable model system with predictable functionality and enhanced ecological stability [2] [3].
In biomedical contexts, SynComs are engineered to model disease-associated microbiomes and develop novel therapeutic interventions. They provide a well-defined, reproducible system to mechanistically study host-microbe interactions, moving beyond correlative observations from complex, variable natural microbiomes [4]. This application note details the design principles, construction protocols, and a specific biomedical application of SynComs for modeling inflammatory bowel disease (IBD).
Computational Design and Metabolic Modeling Frameworks
The rational design of SynComs relies on a Design-Build-Test-Learn (DBTL) cycle, an iterative engineering framework that integrates computational prediction with experimental validation [2] [5]. A critical component of the "Design" phase is comparative metabolic modeling, which predicts the potential for stable coexistence and functional output of candidate strains before laboratory assembly.
Table 1: Key Metrics in Metabolic Modeling for SynCom Design
| Metric | Acronym | Description | Impact on Community |
|---|---|---|---|
| Metabolic Interaction Potential | MIP | Quantifies the potential for cooperative cross-feeding of metabolites [6]. | Higher MIP scores are correlated with increased community stability and cooperation [6]. |
| Metabolic Resource Overlap | MRO | Measures the degree of competition for environmental nutrients and resources [6]. | Lower MRO scores reduce competitive pressure, favoring stable coexistence [6]. |
| Resource Utilization Width | N/A | Reflects the diversity of carbon substrates a strain can metabolize [6]. | Narrow-spectrum utilizers specialize, lowering MRO and increasing MIP, thereby enhancing stability [6]. |
The workflow begins with Genome-Scale Metabolic Models (GEMs), which are computational reconstructions of the metabolic network of an organism. Tools like GapSeq are used to generate these models from genomic data [4]. These individual models are then integrated to simulate community metabolism. Platforms like BacArena enable spatially-resolved, dynamic simulations of microbial communities, modeling nutrient diffusion and cell growth over time to predict whether specific strain combinations can co-exist [4].
Protocols for SynCom Design and Experimental Validation
Protocol 1: Function-Driven Selection and Assembly of SynComs
This protocol outlines the MiMiC2 pipeline for designing a host-specific SynCom based on metagenomic functional profiles [4].
Input Data Preparation:
- Metagenomes: Collect metagenomic sequencing data from the target ecosystem (e.g., gut of healthy vs. diseased individuals).
- Genome Collection: Compile a database of isolated bacterial genomes or high-quality Metagenome-Assembled Genomes (MAGs) from a relevant source (e.g., human gut microbiome).
Functional Annotation:
Function-Based Selection:
- Convert the Pfam annotations for both metagenomes and genomes into binarized presence-absence vectors.
- Assign differential weights to functions:
- Core functions: Identify Pfams present in >50% of the target metagenomes and assign an added weight (e.g., 0.0005) to ensure their inclusion [4].
- Disease-enriched functions: If comparing two groups (e.g., healthy vs. diseased), use a Fisher's exact test to identify Pfams significantly enriched in the target group. Assign these an additional weight (e.g., 0.0012) [4].
- Run the
MiMiC2.pyscript. The algorithm iteratively selects the genome from the collection that best matches the weighted functional profile of the target metagenome, adding it to the SynCom until the desired number of members is reached [4].
In Silico Stability Screening:
Figure 1: Function-Driven SynCom Design Workflow. This diagram outlines the computational pipeline for selecting SynCom members based on metagenomic functional profiles.
Protocol 2: Experimental Validation in a Gnotobiotic Mouse Model
This protocol describes the in vivo testing of a SynCom designed to model a human disease state, specifically Inflammatory Bowel Disease (IBD) [4].
SynCom Cultivation and Formulation:
- Individually culture each bacterial strain in the SynCom under appropriate anaerobic conditions.
- Harvest cells at mid-log phase, centrifuge, and wash with sterile, anaerobic PBS.
- Resuspend all strains and combine them in equal proportions based on cell count (e.g., 10^8 CFU per strain).
Mouse Colonization:
- Use germ-free or antibiotic-pretreated mice (e.g., IL10-/- mice, which are susceptible to colitis) [4].
- Administer the prepared SynCom inoculum to the mice via oral gavage.
- House the mice in gnotobiotic isolators to prevent contamination from other microbes.
Phenotypic Monitoring and Sample Collection:
- Monitor mice daily for signs of clinical disease (e.g., weight loss, piloerection, diarrhea).
- After a pre-defined period (e.g., several weeks), euthanize the mice and collect tissues (colon, cecum contents).
- Process tissues for histological scoring of inflammation and colitis.
Post-Harvest Analysis:
- Microbial Engraftment: Extract DNA from cecal or fecal content. Perform 16S rRNA gene sequencing or shotgun metagenomics to confirm the establishment and stability of the SynCom in vivo.
- Host Response: Analyze cytokine profiles (e.g., by ELISA) and immune cell populations (e.g., by flow cytometry) in colonic tissues to quantify the inflammatory response.
Application Note: An IBD-Mimicking SynCom
Objective: To construct a defined SynCom that recapitulates the functional potential of the human IBD microbiome and induces a colitis phenotype in a susceptible mouse model [4].
SynCom Design:
- Method: The
MiMiC2function-based selection pipeline was applied. - Input: Metagenomic data from ulcerative colitis patients was used as the target ecosystem. A collection of human gut bacterial isolates served as the genome source.
- Key Feature: Functions (Pfam domains) differentially enriched in the IBD metagenomes were assigned higher weights during the selection process, ensuring the SynCom captured the disease-relevant genetic landscape [4].
- Output: A 10-member SynCom (HuSynCom-IBD) was designed.
Experimental Results:
- Germ-free IL10-/- mice were colonized with the HuSynCom-IBD.
- The SynCom successfully stably colonized the mouse gut.
- Mice colonized with HuSynCom-IBD developed significant colitis, as evidenced by weight loss, histopathological scoring of colon sections, and elevated pro-inflammatory cytokine levels, compared to control mice [4].
- This demonstrates the potential of functionally-designed SynComs to model complex human diseases and provide a platform for mechanistic studies and therapeutic screening.
Table 2: Research Reagent Solutions for SynCom Construction & Validation
| Reagent / Material | Function / Application | Example Tools / Strains |
|---|---|---|
| Genome Collections | Source of isolated, sequenced microbes for SynCom assembly. | HiBC (Human), miBC2 (Mouse), Hungate1000 (Rumen) [4] |
| Metabolic Modeling Software | Predicts metabolic interactions and community stability in silico. | GapSeq (model generation), BacArena (dynamic simulation) [4] |
| Function-Based Selection Pipeline | Automates selection of SynCom members from a genome database based on metagenomic functional profiles. | MiMiC2 computational pipeline [4] |
| Gnotobiotic Mouse Model | Provides a sterile, controlled in vivo environment for testing host-SynCom interactions. | IL10-/- mice [4] |
| Pfam Database | Curated database of protein families for functional annotation of genomic and metagenomic data. | Pfam v32 [4] |
Figure 2: Metabolic Principles of SynCom Stability. Narrow-spectrum utilizers specialize, secreting metabolites that others consume, leading to high MIP, low MRO, and stability. Broad-spectrum utilizers compete for the same resources, leading to low MIP, high MRO, and instability.
The rational design of Synthetic Microbial Communities (SynComs) requires a deep integration of core ecological principles with advanced computational modeling. Two foundational concepts—keystone species and metabolic interdependence—provide the theoretical framework for understanding and engineering stable, functional microbial consortia. Keystone species, defined as organisms with disproportionate effects on their environment relative to their abundance [7], play critical roles in maintaining community structure and function. Concurrently, metabolic interdependence describes the complex biochemical network where metabolic byproducts from one organism serve as essential substrates for others within a shared ecosystem [8]. When combined with comparative metabolic modeling, these principles enable researchers to transition from trial-and-error approaches to predictive SynCom design for biomedical, agricultural, and environmental applications [2].
Table 1: Core Ecological Theories and Their Application to SynCom Design
| Ecological Theory | Key Principle | Application in SynCom Design | References |
|---|---|---|---|
| Keystone Species Theory | Species with disproportionate ecological impact | Selection of governance species that enhance community stability and function | [2] [7] |
| Metabolic Interdependence | Cross-feeding of metabolic byproducts | Engineering consortia with complementary nutritional requirements | [8] [9] |
| Metabolic Niche Theory | Organism's metabolic capabilities and requirements | Genome-scale metabolic modeling to predict coexistence | [10] [11] |
| Community Stability Theory | Resistance, resilience, and robustness to perturbation | Designing communities that maintain function under disturbance | [2] |
Computational Protocols for Comparative Metabolic Modeling
Genome-Scale Metabolic Model (GEM) Reconstruction and Analysis
Protocol Objective: Construct and analyze genome-scale metabolic models to predict metabolic capabilities and potential interactions between community members.
Workflow Steps:
- Genome Annotation: Identify metabolic genes and reconstruct metabolic networks from genomic data using tools like ModelSEED, KBase, or RAVEN Toolbox [10] [11]
- Stoichiometric Matrix Construction: Create an ( m \times n ) matrix ( S ) where ( m ) represents internal metabolites and ( n ) represents metabolic reactions [10]
- Constraint Definition: Apply thermodynamic and capacity constraints using the inequality ( lbi \leq vi \leq ubi ), where ( vi ) represents reaction flux [10]
- Steady-State Solution Space: Solve ( Sv = 0 ) to identify all biochemically feasible flux distributions [10]
- Survival Condition Application: Constrain solutions to those satisfying ( v{biomass} \geq v{death} ) to ensure biological viability [10]
Key Computational Metrics:
- Metabolic Interaction Potential (MIP): Quantifies cooperative potential through metabolite exchange [6]
- Metabolic Resource Overlap (MRO): Measures competitive pressure for shared resources [6]
- Niche Breadth: Determines specialization level of metabolic capabilities [10] [11]
Figure 1: Computational workflow for metabolic network reconstruction and analysis
Metabolic Network Analysis for Interaction Prediction
Protocol Objective: Identify potential metabolic interactions and dependencies between community members prior to experimental assembly.
Methodology:
- Complementarity Analysis: Identify pairs of organisms where one secretes metabolites that another requires but cannot synthesize [9]
- Interaction Classification: Categorize relationships as:
- Mutualism: Reciprocal metabolite exchange
- Commensalism: Unidirectional beneficial exchange
- Competition: Overlap in resource requirements [2]
- Keystone Identification: Apply network centrality measures to identify species with disproportionate influence on community metabolic functioning [2] [11]
Table 2: Metabolic Modeling Outputs for SynCom Design Decisions
| Modeling Output | Calculation Method | Design Implication | Stability Impact |
|---|---|---|---|
| Metabolic Interaction Potential (MIP) | Sum of potential cross-feeding interactions | Higher MIP correlates with enhanced cooperation | Positive [6] |
| Metabolic Resource Overlap (MRO) | Measurement of shared nutritional requirements | High MRO indicates competitive pressure | Negative [6] |
| Niche Breadth Index | Diversity of utilizable resources | Narrow-spectrum utilizes enhance complementarity | Positive [6] |
| Interaction Stoichiometry | Quantitative flux of metabolite exchange | Enables optimal ratio determination | Positive [10] |
Experimental Validation Protocols for Designed SynComs
Community Assembly and Stability Assessment
Protocol Objective: Experimentally validate computationally designed SynComs and assess their stability and functional performance.
Workflow Steps:
- Strain Selection: Combine keystone species with narrow-spectrum resource-utilizing bacteria to optimize metabolic interactions while minimizing competition [6]
- Inoculation: Use defined media with controlled nutrient availability to mirror model assumptions
- Longitudinal Monitoring: Track community composition over time (≥20 generations) using 16S rRNA sequencing or strain-specific qPCR [2]
- Functional Assessment: Quantify community-level functions (e.g., biomass production, metabolite secretion) [2]
- Perturbation Response: Evaluate stability through resistance (immediate response) and resilience (recovery capacity) to disturbances [2]
Key Validation Metrics:
- Taxonomic Stability: Maintenance of initial strain ratios over time
- Functional Retention: Preservation of target metabolic activities
- Perturbation Resilience: Return to baseline after environmental stress
Metabolic Interaction Mapping
Protocol Objective: Experimentally verify predicted metabolic interactions and quantify metabolite exchange.
Methodology:
- Spent Media Analysis: Culture individual strains, filter spent media, and test for growth support of partner strains [2]
- Isotope Tracing: Use (^{13})C-labeled metabolites to track cross-feeding relationships [9]
- Spatial Organization: Implement microfluidic devices or agar-based systems to assess distance-dependent interactions [2]
- Metabolomic Profiling: Apply LC-MS/MS to quantify metabolite exchange in co-culture versus monoculture [9]
Figure 2: Experimental validation workflow for synthetic community design
Application Notes: Implementing Ecological Theory in SynCom Design
Environmental Context Integration
The performance of designed SynComs is highly dependent on environmental parameters. Studies of thermophilic communities demonstrate that metabolic interdependencies increase with environmental stress [9]. Under high-temperature conditions (78.5-85.8°C), thermophilic communities exhibited:
- Tighter interaction networks with higher connectivity [9]
- Increased metabolic complementarity, particularly among phylogenetically distant species [9]
- Enhanced archaea-bacteria collaborations as adaptive strategy [9]
These findings highlight the necessity of modeling environmental parameters when designing SynComs for specific applications.
Managing Social Interactions in Microbial Communities
Microbial communities exhibit complex social dynamics that impact stability:
Cheating Behavior Management:
- Problem: Non-producing "cheater" strains exploit public goods without contribution [2]
- Solutions: Spatial structuring, nutrient limitation strategies, and engineering resource dependency [2]
Interaction Balance:
- Optimal communities balance cooperative and competitive interactions [2]
- Excessive cooperation can reduce robustness, while excessive competition destabilizes communities [2]
- Strategic introduction of competitive interactions can enhance stability in certain contexts [2]
Design Principles for Specific Applications
Table 3: Application-Specific SynCom Design Considerations
| Application Domain | Keystone Selection | Metabolic Considerations | Stability Enhancement |
|---|---|---|---|
| Biomedical | Host-adapted commensals with immunomodulatory functions | Host-derived nutrient utilization | Resistance to host defenses and antibiotics |
| Agricultural | Native rhizosphere specialists with plant growth promotion | Root exudate utilization patterns | Resilience to soil perturbations and competition |
| Bioremediation | Pollutant-degrading specialists with complementary pathways | Metabolic division of labor for degradation pathways | Maintenance under fluctuating pollutant loads |
| Industrial Biotechnology | High-yield producers with minimal byproduct formation | Coordinated pathway allocation for target compounds | Stability in bioreactor conditions |
The Scientist's Toolkit: Essential Research Reagents and Platforms
Table 4: Key Research Reagents and Computational Platforms for SynCom Research
| Tool Category | Specific Tools/Platforms | Function | Application Context |
|---|---|---|---|
| Metabolic Modeling Platforms | RAVEN Toolbox, COBRApy, ModelSEED | GEM reconstruction and flux balance analysis | Prediction of metabolic interactions and nutrient requirements [10] |
| Network Analysis Tools | Cytoscape, iNAP, Random Matrix Theory algorithms | Construction and analysis of co-occurrence and metabolic networks | Identification of keystone species and interaction patterns [9] [11] |
| Community Modeling Frameworks | MICOM, SteadyCom, SMET | Multi-species community metabolic modeling | Simulation of cross-feeding and prediction of community stability [2] [12] |
| Experimental Validation Systems | Microfluidic devices, gnotobiotic systems, stable isotope labeling | Controlled testing of predicted interactions | Empirical validation of metabolic dependencies and community dynamics [2] |
| Culture Platforms | High-throughput culturomics, bioreactors | Cultivation of diverse microbial species | Strain isolation and community assembly under controlled conditions [2] |
The integration of keystone species theory with metabolic interdependence concepts provides a powerful framework for designing SynComs with predictable functions and enhanced stability. By employing comparative metabolic modeling as a foundation and validating predictions through rigorous experimental protocols, researchers can advance from empirical community construction to predictive ecosystem engineering. The continued development of computational tools, combined with experimental methods for mapping metabolic interactions, will enable more sophisticated applications across biomedical, agricultural, and environmental domains. Future advances will likely focus on dynamic modeling of community assembly, integration of evolutionary principles, and more sophisticated management of social interactions within engineered consortia.
Understanding the dynamics of microbial interactions—including mutualism, competition, and cheating behavior—is fundamental to advancing synthetic microbial ecology and its applications in biotechnology and medicine. These interactions govern the stability, productivity, and functionality of microbial communities. With the growing emphasis on designing synthetic consortia for industrial processes and therapeutic interventions, the need for precise mapping of these interactions has never been greater. Comparative metabolic modeling using Genome-Scale Metabolic Models (GEMs) provides a powerful computational framework to predict and analyze these complex relationships in silico before embarking on costly experimental work [13]. This Application Note details protocols for integrating GEM-based analysis with experimental validation to systematically map microbial interactions, framed within the broader context of comparative metabolic modeling research for synthetic community engineering.
Key Concepts and Interaction Motifs in Microbial Ecology
Microbial interactions can be categorized into distinct motifs based on their fitness consequences for the involved partners. A clear understanding of this terminology is essential for accurately mapping and interpreting community dynamics.
Table 1: Defining Microbial Interaction Motifs
| Interaction Motif | Description | Impact on Fitness |
|---|---|---|
| Cooperation | An interaction that increases the fitness of neighboring cells. When occurring between cells of the same genotype, it is termed homotypic cooperation [14]. | Beneficial for recipient |
| Mutualism | A cooperative interaction occurring between different genotypes, known as heterotypic cooperation [14]. | Beneficial for both partners |
| Commensalism | An interaction that increases the fitness of a recipient, with no apparent cost or benefit to the donor [14]. | Beneficial for one, neutral for the other |
| Cheating / Parasitism | One member benefits from the interaction at the expense of the donor, or cooperator. This is also known as parasitism [14]. | Beneficial for one, harmful for the other |
| Competition | Both interacting members experience a reduced fitness as a result of their interaction [14]. | Harmful for both partners |
| Amensalism | One partner is negatively affected by the presence of another, which experiences neither cost nor benefit [14]. | Harmful for one, neutral for the other |
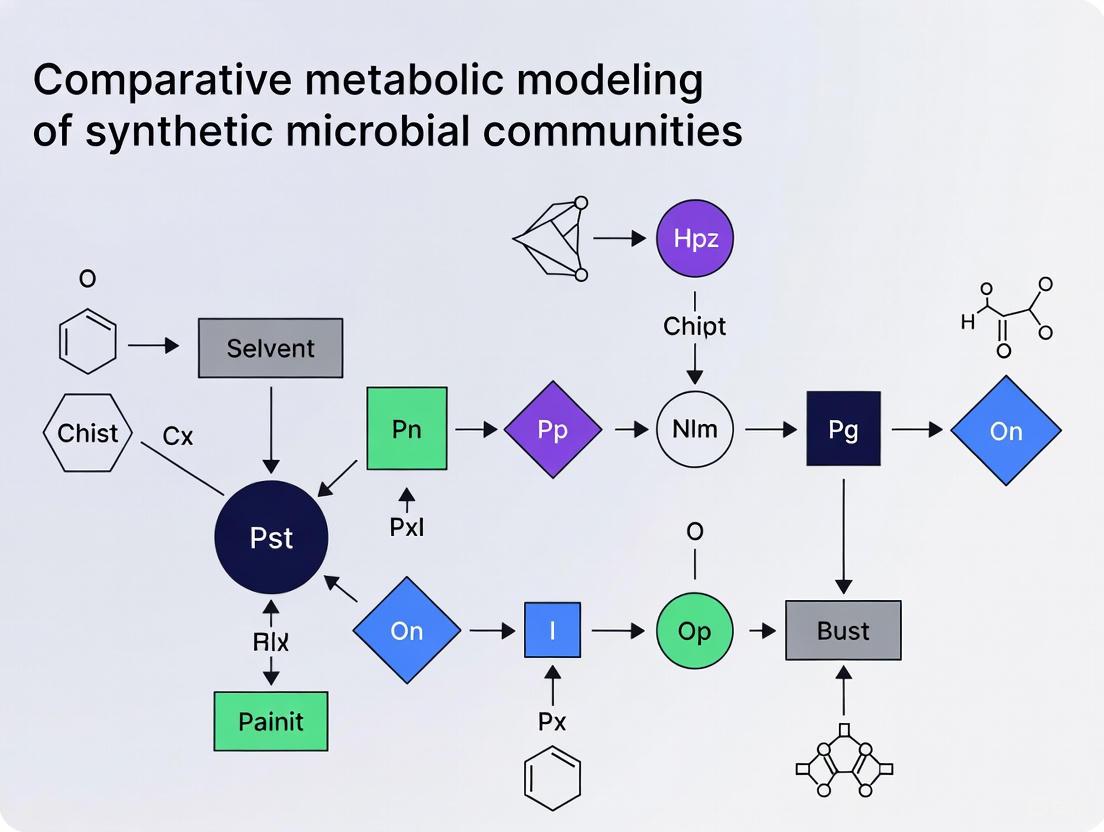
Computational Protocol: Comparative Metabolic Modeling of Interactions
Principle
Genome-Scale Metabolic Models (GEMs) are computational reconstructions of the metabolic network of an organism. They allow for the simulation of metabolic fluxes under given conditions using constraints-based approaches. When applied to communities, GEMs can predict metabolic interactions, such as cross-feeding (a form of mutualism) or competition for resources, by simulating the exchange of metabolites between models [13].
Detailed Workflow for Community GEM Reconstruction and Analysis
Step 1: Model Reconstruction
- Input: Metagenome-Assembled Genomes (MAGs) or isolate genomes.
- Process: Reconstruct individual GEMs using multiple automated tools. Using a consensus approach is critical to minimize tool-specific bias.
- CarveMe: Employs a top-down approach, carving models from a universal template. It is fast and generates models ready for simulation [13].
- gapseq: Uses a bottom-up approach, building models by mapping annotated genomic sequences to reactions. It often yields models with a larger number of reactions and metabolites [13].
- KBase: A bottom-up platform that also utilizes the ModelSEED database for reconstruction [13].
- Output: A set of draft GEMs for each organism in the community.
Step 2: Building a Consensus Community Model
- Rationale: Different reconstruction tools rely on different biochemical databases, leading to variations in the predicted metabolic capabilities and interaction potentials of the models. A consensus approach integrates the outcomes of multiple tools to create a more comprehensive and less biased model [13].
- Process: Merge draft models originating from the same MAG but built with different tools (CarveMe, gapseq, KBase) using a dedicated pipeline [13].
Step 3: Gap-Filling with COMMIT
- Process: Perform gap-filling on the draft consensus community model using the COMMIT tool. This step adds critical reactions to enable growth in a defined medium.
- Use an iterative approach based on MAG abundance (ascending or descending order).
- Initiate with a minimal medium.
- After gap-filling each individual model, predict permeable metabolites and use them to augment the medium for subsequent reconstructions [13].
- Note: The iterative order during gap-filling does not significantly influence the number of added reactions, providing flexibility in the protocol [13].
Step 4: Simulation and Interaction Prediction
- Process: Use constraint-based modeling (e.g., Flux Balance Analysis) on the gap-filled consensus community model.
- Analysis: Identify potential metabolic interactions by analyzing the flux through exchange reactions. This helps predict cross-fed metabolites (mutualism/commensalism) and metabolites competed for (competition).
The following workflow diagram outlines this multi-step computational protocol:
Experimental Protocol: Validating Predicted Interactions
Principle
Computational predictions of microbial interactions, such as mutualistic cross-feeding or cheating, require experimental validation. This protocol uses engineered microbial strains to verify and quantify these interactions in controlled laboratory environments.
Detailed Workflow for Validating Cross-Feeding Mutualism
Step 1: Engineer Mutualistic Strains
- Objective: Create two (or more) microbial strains whose growth depends on metabolites exchanged between them.
- Procedure:
- Example System: Engineer two auxotrophic yeast strains, such as:
- Genetic Modifications: Use gene knockouts to create the auxotrophies and promoter engineering to enhance the production and secretion of the complementary metabolite.
Step 2: Co-culture and Monitor Population Dynamics
- Setup: Inoculate the engineered strains together in a minimal medium that lacks both essential metabolites (leucine and tryptophan in this example). Neither strain can grow alone in this medium.
- Monitoring: Measure the population dynamics of each strain over time using:
- Flow Cytometry: To obtain absolute cell counts and assess co-culture stability [15].
- Optical Density (OD): For general growth monitoring.
- Plating and Colony Counting: On selective media to distinguish strains.
Step 3: Quantify Interaction Strength and Identify Cheaters
- Testing for Stability: Co-culture the mutualistic strains with a potential "cheater" strain. A cheater is engineered to be unable to produce the required public good (e.g., an invertase-deficient yeast strain in a sucrose medium [14]) but can still consume the metabolites produced by the cooperators.
- Analysis: Track the frequency of cooperators and cheaters over multiple growth cycles. A stable mutualism will resist invasion by the cheater, often through mechanisms like preferential access to the exchanged nutrients [14].
The experimental workflow for this validation is depicted below:
The Scientist's Toolkit: Research Reagent Solutions
Table 2: Essential Reagents and Tools for Mapping Microbial Interactions
| Reagent / Tool | Function / Application | Key Considerations |
|---|---|---|
| CarveMe [13] | Automated top-down reconstruction of Genome-Scale Metabolic Models (GEMs). | Fast; uses a universal template. May produce models with fewer reactions than bottom-up tools. |
| gapseq [13] | Automated bottom-up reconstruction of GEMs from annotated genomes. | Can produce more comprehensive models; uses multiple data sources. May generate more dead-end metabolites. |
| KBase [13] | Integrated platform for bottom-up GEM reconstruction and community analysis. | User-friendly; uses ModelSEED database. Results may be similar to gapseq due to shared database. |
| COMMIT [13] | A tool for gap-filling metabolic models in a community context. | Iteratively updates the medium based on secreted metabolites; order of gap-filling has minimal impact on results. |
| Flow Cytometry [15] | Quantifies absolute microbial cell counts in a sample for QMP. | Counts only intact cells, ignoring free extracellular DNA. Essential for normalizing sequencing data to absolute abundance. |
| Propidium Monoazide (PMA) [15] | Treatment to remove DNA from dead/membrane-compromised cells before DNA extraction. | Helps focus analysis on the intact/viable microbiome. May not fully reconcile differences between cell-counting and DNA-based quantification. |
| qPCR / ddPCR [15] | Molecular methods to quantify total microbial load by targeting the 16S rRNA gene. | Cost-effective and accessible (qPCR). Digital Droplet PCR (ddPCR) offers greater precision and sensitivity. |
Application in Drug Development: Pharmacomicrobiomics and Pharmacoecology
The mapping of microbial interactions has direct relevance for drug development, particularly in the emerging fields of pharmacomicrobiomics and pharmacoecology.
- Pharmacomicrobiomics studies how the microbiome influences drug distribution, metabolism, efficacy, and toxicity [16] [17]. For instance, gut microbes can directly biotransform drugs (e.g., the cardiac drug digoxin by Eggerthella lenta) or bioaccumulate them, thereby altering drug availability and activity [17].
- Pharmacoecology describes the impact of drugs, including non-antibiotics, on the composition and function of the microbiome [17]. Many drugs, from antidiabetics to proton pump inhibitors, have been shown to exert off-target antimicrobial effects, altering the microbial ecology [17].
Understanding these bidirectional interactions is critical for explaining Individual Variability in Drug Response (IVDR) and for designing personalized therapeutic strategies that account for an individual's microbiome composition [16]. The protocols outlined in this document for mapping interactions can be applied to study how drugs modulate microbial community dynamics (pharmacoecology) and how these changes, in turn, affect drug metabolism and efficacy (pharmacomicrobiomics).
A fundamental paradigm in microbial ecology is that the behavior of a consortium is not a simple, linear sum of the behaviors of its individual members. This is the core of nonlinear scaling, where emergent properties arise from the complex web of interactions between organisms in a defined community. For research focused on the comparative metabolic modeling of synthetic microbial communities (SynComs), recognizing, quantifying, and predicting this nonlinearity is paramount [2]. The shift from empirical community construction to predictive ecosystem engineering relies on a mechanistic understanding of these interactions [2]. Defined in vitro communities provide a tractable system to dissect these complexities, offering a bridge between simplistic monoculture studies and the overwhelming intricacy of natural microbiomes [18]. This Application Note outlines the theoretical frameworks, quantitative methodologies, and practical protocols essential for investigating nonlinear scaling in SynComs.
Theoretical Foundations of Nonlinear Interactions
Nonlinearity in SynComs primarily stems from the dynamic and context-dependent nature of microbial interactions. These can be categorized and modeled to inform experimental design.
Ecological Interaction Types and Their Consequences
Microbial interactions define the stability and function of a consortium. The major types of interactions include:
- Positive Interactions (Mutualism & Commensalism): Often emerge from metabolic cross-feeding, where the exchange of metabolic byproducts enhances overall community efficiency and resilience [2]. For instance, engineered cross-feeding yeast consortia have demonstrated increased production of target compounds like 3-hydroxypropionic acid [2].
- Negative Interactions (Competition & Antagonism): These arise from competition for limited resources (nutrients, space) or through chemical warfare via antimicrobial compounds (e.g., antibiotics, bacteriocins) [2]. The outcome of competition can be strongly predicted by phylogenetic relatedness and the overlap of biosynthetic gene clusters [2].
- Cheating Behavior: A significant challenge for community stability, cheating occurs when some members exploit public goods without contributing, potentially leading to the collapse of mutualistic partnerships [2]. Spatial structuring of communities is a key strategy to mitigate cheating by altering quorum sensing dynamics and public goods distribution [2].
Quantifying Interaction Dynamics with Flow Cytometry
A significant bottleneck in SynCom research is the rapid quantification of individual taxon abundances. Flow cytometry (FC), combined with supervised classification, presents a high-throughput solution. This method involves training a classifier on FC data from monocultures and applying it to assign cells in mixed communities to specific species, providing species-specific cell counts [19]. It performs equally well or better than 16S rRNA gene sequencing for quantifying species in defined cocultures and avoids biases from varying gene copy numbers and amplification efficiencies [19].
Table 1: Key Experimental Models for Studying Defined Microbial Communities
| Model System | Description | Key Applications | Considerations |
|---|---|---|---|
| Gnotobiotic Mice | Germ-free animals colonized with a defined microbial consortium [18]. | Studying host-microbe interactions, immune response, and pathogen resistance in a whole-organism context [18]. | Limited translational fidelity to humans; high operational costs [18]. |
| In Vitro Cocultures | Defined communities cultivated in controlled laboratory media [19]. | Unraveling fundamental microbe-microbe interactions, metabolic cross-talk, and community assembly rules [2]. | Lacks host factors; may oversimplify complex natural environments. |
| Gut-on-a-Chip / Organoids | Sophisticated in vitro models mimicking human intestinal physiology [18]. | Investigating host-microbe interactions with more human relevance than animal models [18]. | Technologically complex; may not fully capture systemic host responses. |
Quantitative Data on Nonlinear Outcomes
The following data, derived from empirical studies, exemplifies the nonlinear dynamics of SynComs.
Table 2: Manifestations of Nonlinear Scaling in Synthetic Microbial Communities
| Nonlinear Phenomenon | Experimental Context | Observed Outcome | Implication for SynCom Design |
|---|---|---|---|
| Interaction Shift | Chlorella vulgaris-Saccharomyces cerevisiae consortium under elevated NH₄⁺ [2]. | Transition from mutualism to competition. | Abiotic conditions (nutrient levels) can fundamentally alter interaction types. |
| Emergent Competition | Three-member cross-feeding SynCom upon introduction of a fourth strain [2]. | Reduction in the yield of the target compound, 4-ethylclove acid. | Community expansion can trigger unforeseen competitive interactions that reduce function. |
| Cheater Exploitation | SynComs based on public goods production (e.g., siderophores, enzymes) [2]. | Collapse of cooperative partnerships and loss of community function. | Stability requires engineering strategies to suppress cheating, such as spatial structuring. |
| Keystone Species Effect | Introduction or removal of a keystone species from a community [2]. | Disproportionate impact on community structure, stability, and functional output. | Identification and inclusion of keystone taxa are critical for consortium robustness. |
Experimental Protocols
This section provides a detailed methodology for a key experiment investigating nonlinear growth dynamics in a defined coculture.
Protocol: Quantifying Species Abundance in Cocultures using Flow Cytometry and Supervised Classification
Objective: To accurately quantify the relative abundance of individual bacterial species in a defined coculture over time, enabling the analysis of nonlinear population dynamics.
Materials:
- Bacterial Strains: Selected human gut bacteria (e.g., Bacteroides thetaiotaomicron, Faecalibacterium prausnitzii, Collinsella aerofaciens).
- Growth Media: Modified Gifu Anaerobic Medium (mGAM) broth, Reinforced Clostridial Medium (RCM) broth.
- Equipment: Anaerobic workstation (10% H₂, 10% CO₂, 80% N₂), flow cytometer (e.g., BD Accuri C6 or CytoFLEX S), plate reader.
- Reagents: SYBR Green I nucleic acid stain, phosphate-buffered saline (PBS), dimethylsulfoxide (DMSO), validation beads for flow cytometer calibration.
Procedure:
- Monoculture Preparation and Standardization:
- Subculture each bacterial strain twice under anaerobic conditions at 37°C until stationary phase is reached (confirmed by OD₆₀₀ monitoring) [19].
- Dilute each monoculture 1000-fold in PBS to a density of ~10⁶ cells/mL.
- Stain cells with 1 µL/mL SYBR Green I (diluted 1:100 in DMSO) and incubate at 37°C for 20 minutes [19].
Flow Cytometry Data Acquisition for Training Set:
- Calibrate the flow cytometer daily using validation beads.
- For each monoculture, acquire multiparametric FC data (FSC, SSC, FL1-H, etc.) for at least 10,000 events [19]. This data forms the training set for the supervised classifier.
Construction of In Vitro Mock Communities:
- Based on cell density counts from the flow cytometer, mix the standardized monoculture suspensions in a series of intended proportions (e.g., 5%, 10%, 20%, 40%, 50%, 60%, 80%, 90%, 95%) in a final volume of 1 mL of filtered PBS [19]. Note that the expected proportions will differ slightly from the intended ones due to pipetting errors.
Classifier Training and Validation:
- Use a supervised classification algorithm (e.g., Random Forest, Linear Discriminant Analysis) trained on the monoculture FC data.
- Apply the trained classifier to the FC data from the mock communities of known expected proportions to validate its prediction accuracy [19].
Co-growth Community Experiment:
- Inoculate a fresh culture medium (e.g., RCM) with multiple species in equal proportions to a total concentration of ~4x10⁶ cells/mL [19].
- Incubate the community under anaerobic conditions at 37°C.
- Sample the community at designated timepoints (e.g., 24h, 48h).
- For each sample, perform FC analysis and use the trained classifier to predict the relative abundance of each species.
Data Analysis:
- Compare the classifier-predicted proportions against the expected proportions for validation.
- For the co-growth experiment, plot the population dynamics of each species over time to identify nonlinear behaviors, such as competitive exclusion or cooperative growth.
Visualization of Concepts and Workflows
The following diagrams, defined using the DOT language, illustrate the core concepts and experimental workflows.
Diagram 1: Nonlinear Interaction Network in a SynCom
Title: Nonlinear Interaction Network in a SynCom
Diagram 2: Flow Cytometry Quantification Workflow
Title: Flow Cytometry Species Quantification Workflow
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential Research Reagents and Resources for SynCom Studies
| Reagent / Resource | Function / Description | Key Consideration |
|---|---|---|
| Defined Microbial Strains | Individual, well-characterized bacterial isolates from culture collections (e.g., DSMZ, ATCC) or human feces [19]. | Genomic and metabolic characterization is crucial for interpreting interaction data. |
| Gnotobiotic Animal Models | Germ-free mice or rats for in vivo host-microbe interaction studies [18]. | The Altered Schaedler Flora (ASF) is a classic defined consortium for standardizing mouse microbiota [18]. |
| Genome-Scale Metabolic Models (GEMs) | Computational models that predict organism metabolism; can be extended to microbial communities [20]. | Enable in silico simulation of metabolic interactions and resource partitioning within SynComs [20]. |
| Anaerobic Culture Systems | Workstations or chambers providing an oxygen-free atmosphere (e.g., 10% H₂, 10% CO₂, 80% N₂) for cultivating obligate anaerobes [19]. | Essential for maintaining the viability of many gut-derived bacterial species. |
| Flow Cytometry with Supervised Classification | High-throughput, single-cell analysis for quantifying species abundances in a community without sequencing [19]. | Performance is species-dependent; requires training a classifier on monoculture data first [19]. |
Application Notes
This document provides a standardized framework for quantifying and evaluating the stability, robustness, and functional resilience of Synthetic Microbial Communities (SynComs). These metrics are vital for transitioning SynComs from controlled laboratory settings into predictable applications in biotechnology, medicine, and agriculture.
Quantitative Metrics for SynCom Performance
The following table summarizes the core quantitative metrics used to assess SynCom stability and function, derived from recent experimental studies.
Table 1: Key Quantitative Metrics for Assessing SynCom Stability and Resilience
| Metric Category | Specific Metric | Measurement Method | Reported Value | Context |
|---|---|---|---|---|
| Functional Stability | Denitrification Efficiency | NO3−-N removal rate [21] | Maintained at ~93% | Under disturbances from Dibutyl Phthalate (DBP) and Levofloxacin (LOFX) [21] |
| Compositional Resilience | Abundance of Persistent Strains | Flow cytometry (Live/Dead cell counts) [22] | 81% reduction in live cells | For a persistent Pseudomonas strain exposed to native soil microbes [22] |
| Structural Stability | Metabolic Resource Overlap (MRO) | Genome-scale Metabolic Modeling (GMM) [6] | Lower values correlate with higher stability | Negative correlation with community stability [6] |
| Structural Stability | Metabolic Interaction Potential (MIP) | Genome-scale Metabolic Modeling (GMM) [6] | Higher values correlate with higher stability | Positive correlation with community stability [6] |
| Functional Output | Plant Dry Weight Increase | Biomass measurement [6] | >80% increase | For stable SynComs (SynCom4 & SynCom5) in the tomato rhizosphere [6] |
Mechanistic Insights into Stability
Advanced omics technologies have elucidated key molecular and ecological mechanisms underpinning SynCom resilience:
- Interspecific Division of Labor: Under environmental disturbances, community members dynamically reallocate functional roles. For instance, in an aerobic denitrification SynCom, Dibutyl Phthalate (DBP) disturbance stimulated Aeromonas hydrophila and Pseudomonas aeruginosa to become functionally dominant, whereas Levofloxacin (LOFX) induced Acinetobacter baumannii and P. aeruginosa to play major roles [21].
- Molecular Cross-Talk: Quorum sensing (QS) via signaling molecules like N-butyryl-L-homoserine lactone (C4-HSL) and N-(3-oxododecanoyl)-L-homoserine lactone (3OC12-HSL) is a critical mechanism for coordinating community responses to stress, enhancing microbial activity, and facilitating adaptation [21].
- Metabolic Network Reprogramming: Stability is maintained through the redirection of electron and energy fluxes. Disturbances can trigger the acceleration of the Tricarboxylic Acid (TCA) cycle, boost electron transfer activity, and upregulate denitrification enzyme expression, ensuring core functions proceed unimpeded [21].
- Niche Specialization: Strains with narrow-spectrum resource utilization (NSR) profiles demonstrate a reduced Metabolic Resource Overlap (MRO) and increased Metabolic Interaction Potential (MIP), which significantly enhances community stability by minimizing competition and fostering obligate cross-feeding interactions [6].
Experimental Protocols
Protocol 1: Assessing Functional Resilience to Chemical Perturbations
This protocol details a method for evaluating the stability of a SynCom's metabolic function when exposed to environmental contaminants, adapted from a study on aerobic denitrification [21].
1. Objectives:
- To determine the resilience of a specific SynCom function (e.g., denitrification) under chemical stress.
- To quantify key physiological and molecular responses that underpin functional stability.
2. Materials:
- SynCom Members: Pre-cultured strains (e.g., Pseudomonas aeruginosa, Acinetobacter baumannii, Aeromonas hydrophila).
- Basal Medium: Defined mineral medium.
- Target Compound: Sodium Nitrate (NO3−-N).
- Chemical Stressors: e.g., Dibutyl Phthalate (DBP), Levofloxacin (LOFX).
- Analytical Equipment: HPLC, LC-MS/MS.
- Reagent Kits: For EPS extraction, DNA/RNA extraction.
3. Procedure:
- Step 1: System Start-up. Inoculate the SynCom into a semi-continuous bioreactor containing the basal medium and the target compound. Operate until a stable functional performance is achieved (e.g., >94% denitrification efficiency).
- Step 2: Introduction of Disturbance. Once stable, introduce the chemical stressor at an environmentally relevant concentration. Maintain a control reactor without the stressor.
- Step 3: Functional Monitoring. Regularly monitor the key functional output (e.g., NO3−-N concentration) to calculate removal efficiency.
- Step 4: Molecular & Physiological Analysis. At key time points, sample the community for:
- Quorum Sensing Molecules: Quantify AHL types (e.g., C4-HSL, 3OC12-HSL) using LC-MS/MS [21].
- Electron Transfer Activity: Measure cytochrome c content and electron transfer system activity.
- Metabolomics/Transcriptomics: Analyze changes in the TCA cycle and denitrification pathway genes via metatranscriptomics.
- Step 5: Data Analysis. Correlate functional performance data with molecular data to establish mechanisms of resilience.
Protocol 2: Quantifying Compositional Resilience Against Native Microbiomes
This protocol assesses the ability of a SynCom to persist and maintain its composition when challenged by a complex native soil microbiome [22].
1. Objectives:
- To measure the survival and persistence of individual SynCom strains in a biotic stress environment.
- To identify strains with inherent "persistent traits."
2. Materials:
- SynCom: Defined consortium of strains (e.g., six compatible Pseudomonas species).
- Native Soil Microbiome: Fresh soil suspension.
- Experimental Hardware: Transwell system with permeable membranes.
- Growth Medium: Appropriate defined medium.
- Flow Cytometer: For viability counts.
3. Procedure:
- Step 1: Experimental Setup. Place the SynCom in one compartment of the transwell system and the native soil microbiome in the other. This allows chemical cross-talk but prevents physical contact.
- Step 2: Incubation and Sampling. Incubate the system and sample both compartments at regular intervals over a defined period.
- Step 3: Viability Analysis. Use flow cytometry with live/dead staining to quantify the abundance and viability of each SynCom strain.
- Step 4: Phenotypic Profiling (Optional). For persistent strains, profile their metabolic utilization of key compound classes (polymers, carboxylic acids, amino acids, etc.) using phenotype microarrays to understand their metabolic strategy under stress [22].
- Step 5: Data Analysis. Calculate the percentage reduction in live cells for each strain. Strains showing significantly higher persistence can be identified as core contributors to resilience.
Protocol 3: Computational Prediction of Community Stability Using Metabolic Modeling
This protocol leverages Genome-scale Metabolic Models (GMMs) to predict the intrinsic stability of a SynCom during the design phase [6].
1. Objectives:
- To computationally screen candidate SynComs for high stability.
- To identify strains that contribute most to cooperative interactions.
2. Materials:
- Genomic Data: Annotated genome sequences for all candidate strains.
- Phenotypic Data (Optional): Phenotype microarray data on carbon source utilization.
- Software: Metabolic modeling tools (e.g., RAVEN, COBRA, CarveMe).
- Computing Environment: Standard workstation or high-performance computing cluster.
3. Procedure:
- Step 1: Model Reconstruction. Build draft Genome-scale Metabolic Models (GEMs) for each candidate strain using an automated pipeline.
- Step 2: Model Refinement. Refine the draft models using phenotypic data (e.g., from Biolog assays) to improve context-specific accuracy [6].
- Step 3: Community Simulation. Simulate all possible combinations of the candidate strains (pairwise and higher-order) using constraint-based methods like Flux Balance Analysis (FBA).
- Step 4: Metric Calculation. For each simulated community, calculate two key indices:
- Metabolic Resource Overlap (MRO): An indicator of competition.
- Metabolic Interaction Potential (MIP): An indicator of cooperation [6].
- Step 5: In Silico Selection. Prioritize community designs that exhibit low MRO and high MIP for experimental validation.
The Scientist's Toolkit
Table 2: Essential Research Reagent Solutions for SynCom Stability Research
| Item Name | Function/Application | Specific Example |
|---|---|---|
| AHL Standards | Quantification of quorum sensing signals via LC-MS/MS to monitor interspecies communication. | C4-HSL, 3OC12-HSL [21] |
| Phenotype Microarrays | High-throughput profiling of carbon source utilization to determine resource utilization width and overlap. | Biolog plates [6] |
| Transwell Co-culture Systems | Physically separate but chemically connect SynComs and native microbiomes to study biotic resilience. | Permeable membrane inserts [22] |
| Genome-scale Metabolic Modeling (GMM) Software | In silico prediction of metabolic interactions, MRO, and MIP to guide stable community design. | RAVEN, COBRA, ModelSEED [23] [6] |
| Semi-continuous Bioreactors | Maintain SynComs in a steady state for long-term functional stability studies under perturbation. | Lab-scale fermenters [21] |
Visualizations
SynCom Stability Analysis Workflow
Mechanisms of Functional Resilience
A Toolkit for Prediction: Metabolic Modeling, DBTL Cycles, and Data-Driven Design
Genome-Scale Metabolic Models (GEMs) as Predictive Blueprints
Genome-scale metabolic models (GEMs) are sophisticated computational tools that enable the mathematical simulation of metabolism across all domains of life, including archaea, bacteria, and eukaryotic organisms [24]. These models quantitatively define the relationship between genotype and phenotype by integrating various types of big data, including genomics, metabolomics, and transcriptomics [24]. GEMs represent structured knowledge-bases that abstract critical information on the biochemical transformations within specific target organisms, containing all known metabolic information including genes, enzymes, reactions, associated gene-protein-reaction (GPR) rules, and metabolites [24].
The reconstruction and application of GEMs have become standard systems biology approaches for modeling cellular physiology and growth, with extensions of this methodology emerging as valuable avenues for predicting, understanding, and designing microbial communities [25]. By converting reconstructions into mathematical formats, researchers can conduct myriad computational biological studies, including network content evaluation, hypothesis testing and generation, analysis of phenotypic characteristics, and metabolic engineering [26]. The capacity to simulate metabolic behavior in silico makes GEMs particularly powerful for both basic research and applied biotechnology.
Reconstruction Protocols and Methodologies
Comprehensive Reconstruction Workflow
The process of building high-quality genome-scale metabolic reconstructions follows a detailed protocol encompassing several critical stages [26]. This structured approach ensures the production of quality-controlled, quality-assured (QC/QA) reconstructions that maintain high standards and comparability between different models. The reconstruction process typically requires significant time investment, ranging from six months for well-studied, medium genome-sized bacteria to two years for complex reconstructions such as human metabolism [26].
Table 1: Key Stages in Metabolic Network Reconstruction
| Stage | Description | Primary Outputs |
|---|---|---|
| Stage 1: Draft Reconstruction | Initial compilation of metabolic genes, reactions, and metabolites from genomic and biochemical databases | Draft metabolic network |
| Stage 2: Manual Refinement | Curation of network content, including organism-specific features and reaction directionality | Curated metabolic reconstruction |
| Stage 3: Conversion to Mathematical Model | Implementation of constraint-based modeling framework and definition of objective functions | Stoichiometric matrix and model constraints |
| Stage 4: Network Validation | Debugging and verification of model functionality against experimental data | Validated, functional metabolic model |
| Stage 5: Application | Utilization for hypothesis testing, experimental design, and prediction | Model predictions and biological insights |
The reconstruction process begins with creating a draft reconstruction from genomic data, followed by manual refinement to incorporate organism-specific biochemical knowledge [26]. This draft is subsequently converted into a mathematical model suitable for constraint-based analysis, validated through debugging procedures, and finally applied to address specific biological questions. Throughout this process, the reconstruction acts as a biochemical, genetic, and genomic (BiGG) knowledge-base for the target organism [26].
Tools and Databases for Reconstruction
Numerous software tools and databases support the reconstruction process, each offering distinct capabilities and relying on different biochemical databases that can significantly influence the resulting models [27]. A comparative analysis of reconstruction tools reveals that CarveMe, gapseq, and KBase represent three prominent automated approaches, each with unique characteristics and advantages.
Table 2: Comparison of Automated GEM Reconstruction Tools
| Tool | Reconstruction Approach | Primary Database | Key Features | Model Characteristics |
|---|---|---|---|---|
| CarveMe | Top-down | Universal template | Fast model generation | Highest number of genes |
| gapseq | Bottom-up | Multiple comprehensive sources | Extensive biochemical information | Most reactions and metabolites |
| KBase | Bottom-up | ModelSEED | User-friendly platform | Intermediate gene count |
| Consensus | Hybrid | Combined sources | Reduced dead-end metabolites | Comprehensive reaction coverage |
The selection of reconstruction tools significantly impacts model structure and predictive capacity. Studies have demonstrated that despite being reconstructed from the same metagenome-assembled genomes (MAGs), different approaches yield markedly different results [27]. For instance, gapseq models typically encompass more reactions and metabolites, while CarveMe models contain the highest number of genes [27]. Consensus models, formed by integrating reconstructions from multiple tools, have shown promise in reducing uncertainty and improving functional capability by retaining majority unique reactions and metabolites while reducing dead-end metabolites [27].
GEM Applications in Microbial Community Modeling
Modeling Synthetic Microbial Communities
Genome-scale metabolic modeling of microbial communities represents a powerful extension of single-organism modeling, enabling investigation of metabolic interactions and community-level functionalities [28] [25]. The Computation of Microbial Ecosystems in Time and Space (COMETS) platform extends dynamic flux balance analysis to simulate multiple microbial species in molecularly complex and spatially structured environments [25]. This approach incorporates accurate biophysical modeling of microbial biomass expansion, evolutionary dynamics, and extracellular enzyme activity modules, providing a comprehensive framework for simulating community behaviors.
Several approaches exist for constructing community-scale metabolic models, each suited to different research objectives. The "mixed-bag" approach integrates all metabolic pathways into a single model with one cytosolic and one extracellular compartment, suitable for analyzing interactions between communities [27]. Compartmentalization combines multiple GEMs into a single stoichiometric matrix with distinct compartments for each species, while costless secretion employs dynamically updated media based on exchange reactions [27]. The choice of methodology depends on the specific research questions and community characteristics being investigated.
Investigating Higher-Order Microbial Interactions
Recent research has employed GEMs to investigate emergent metabolic behaviors in controlled synthetic communities of varying complexity [28]. A 2025 study analyzed synthetic anaerobic communities containing two, three, or four species representing core metabolic guilds in cellulose degradation and carbon conversion [28]. The researchers applied a systems biology framework combining proteogenomics, stoichiometric flux modeling, and Species Metabolic Coupling Analysis (SMETANA) to quantify syntrophic cooperation and competition across configurations.
This research revealed that microbial cooperation peaks in tri-cultures and declines nonlinearly in more complex assemblies, demonstrating that interaction strength depends more on metabolic compatibility than mere species richness [28]. The study documented context-dependent functional roles, with Ruminiclostridium cellulolyticum serving as the dominant metabolite donor while adjusting its enzyme expression based on partner identity, and Methanosaeta concilii becoming fully metabolite-dependent while enhancing methanogenesis [28]. These findings illustrate how GEMs can resolve metabolic network rewiring across defined communities, providing a framework for interpreting and engineering stable, functionally interdependent microbial ecosystems.
Figure 1: Workflow for Metabolic Modeling of Microbial Communities
Metabolic Modeling for Metabolic Profile Predictions
Predicting Biomarkers and Metabolic Perturbations
GEMs have been successfully applied to predict metabolic profiles resulting from genetic variations or disease states [29]. The SAMBA (SAMpling Biomarker Analysis) approach exemplifies this application by simulating fluxes in exchange reactions following metabolic perturbations using random sampling [29]. This method compares simulated flux distributions between baseline and modulated conditions, ranking predicted differentially exchanged metabolites as potential biomarkers for specific perturbations.
This computational approach assists in experimental design by predicting which metabolites are most likely to show differential abundance under given metabolic conditions, thereby guiding resource-intensive metabolomics studies [29]. Validation studies have demonstrated good concordance between simulated metabolic exchange profiles and experimental differential metabolites detected in plasma, including patient data from disease databases and metabolic trait-SNP associations from genome-wide association studies [29]. This capability enables researchers to prioritize metabolites for experimental analysis and gain insights into underlying metabolic pathway perturbations.
Integrating Machine Learning with Constraint-Based Modeling
The integration of machine learning with constraint-based modeling represents an emerging frontier in metabolic modeling research [30]. Although this integration is still in its early stages, it holds significant promise for enhancing both model parameterization and biological insight generation. Machine learning approaches can identify meaningful features from large-scale data and connect them to biological mechanisms, helping establish causality in genotype-phenotype relationships [30].
Iterative integrative schemes represent a particularly promising approach, where machine learning fine-tunes input constraints in constraint-based models [30]. Conversely, constraint-based model simulation results can be analyzed by machine learning and reconciled with experimental data, creating refinement cycles that continue until consistency is achieved between experimental data, machine learning results, and model simulations [30]. This synergistic approach has the potential to enhance both predictive accuracy and mechanistic understanding of metabolic systems.
Experimental Protocols and Methodologies
Protocol for Community Model Reconstruction and Simulation
The reconstruction of community metabolic models follows a systematic protocol that builds upon established single-species methodologies while incorporating community-specific considerations:
Draft Reconstruction: Generate individual GEMs for all community members using automated tools (CarveMe, gapseq, or KBase) or manual curation [26] [27]. Consensus approaches that integrate multiple reconstruction tools may reduce uncertainty and improve model quality [27].
Model Integration: Combine individual GEMs using compartmentalization, mixed-bag, or other appropriate approaches based on research objectives [27]. Standardize metabolite and reaction namespaces to ensure compatibility between models.
Gap-Filling: Implement an iterative gap-filling process using tools such as COMMIT, initiating with a minimal medium and dynamically updating permeable metabolites after each model's gap-filling step [27]. Studies indicate that the iterative order during this process does not significantly influence the number of added reactions [27].
Constraint Definition: Define appropriate physiological and environmental constraints, including nutrient availability, thermodynamic considerations, and spatial parameters when using platforms like COMETS [25].
Model Validation: Compare simulation results with experimental data on community composition, metabolic exchanges, and functional outputs to assess model predictive capability [26].
Simulation and Analysis: Implement appropriate simulation techniques (e.g., dynamic FBA, COMETS) to investigate community metabolic behaviors and interaction patterns [28] [25].
Table 3: Essential Resources for GEM Reconstruction and Analysis
| Category | Resource | Function | Application Context |
|---|---|---|---|
| Genome Databases | Comprehensive Microbial Resource (CMR) | Provides annotated genomic data | Draft reconstruction |
| Genomes OnLine Database (GOLD) | Catalog of genome projects | Genome availability assessment | |
| NCBI Entrez Gene | Gene-centered information | Gene function annotation | |
| Biochemical Databases | KEGG | Metabolic pathway information | Reaction and pathway annotation |
| BRENDA | Enzyme functional data | Enzyme characterization | |
| Transport DB | Membrane transport data | Transport reaction annotation | |
| Modeling Software | COBRA Toolbox | Constraint-based reconstruction and analysis | Model simulation and analysis |
| COMETS | Microbial ecosystem simulation | Spatiotemporal community modeling | |
| CarveMe | Automated model reconstruction | Rapid GEM generation | |
| MEMOTE | Model testing | Quality assessment | |
| Analysis Tools | SMETANA | Species Metabolic Coupling Analysis | Metabolic interaction quantification |
| SAMBA | Sampling Biomarker Analysis | Metabolic biomarker prediction |
Figure 2: GEM Development and Validation Workflow
Genome-scale metabolic models serve as predictive blueprints that enable researchers to simulate and analyze metabolic capabilities across individual organisms and complex microbial communities. The continued refinement of reconstruction methodologies, including the development of consensus approaches and integration of machine learning, enhances model predictive accuracy and biological relevance. As these tools become increasingly sophisticated and accessible, they promise to deepen our understanding of microbial interactions and enable more effective engineering of microbial communities for biomedical, biotechnological, and environmental applications. The structured protocols and resources outlined in this article provide researchers with essential guidance for leveraging GEMs in comparative metabolic modeling of synthetic microbial communities.
The Design-Build-Test-Learn (DBTL) Cycle for Iterative Community Optimization
Application Note
The Design-Build-Test-Learn (DBTL) cycle provides a powerful, iterative framework for optimizing Synthetic Microbial Communities (SynComs), enabling the transition from trial-and-error approaches to predictable ecosystem engineering [2]. This structured process is particularly crucial for overcoming functional instability in applied communities, a challenge stemming from our incomplete understanding of intricate microbial dynamics [6]. By integrating computational modeling, high-throughput experimentation, and data-driven learning, the DBTL cycle allows researchers to systematically optimize community composition for enhanced stability, functionality, and resilience in target environments such as the rhizosphere, gut, or bioreactors [2] [6].
The core innovation within modern DBTL cycles lies in the strategic incorporation of ecological principles and comparative metabolic modeling during the Design phase, and the application of machine learning in the Learn phase to extract meaningful patterns from complex data [31] [2]. This approach is exemplified by recent research demonstrating that narrow-spectrum resource-utilizing bacteria, such as Cellulosimicrobium cellulans E and Pseudomonas stutzeri G, significantly enhance community stability by increasing metabolic interaction potential and reducing metabolic resource overlap [6]. The iterative nature of the DBTL cycle allows for the refinement of these ecological hypotheses, ultimately leading to the construction of SynComs with predictable and robust behaviors for applications in agriculture, biomedicine, and environmental remediation [2].
Experimental Protocols
Protocol 1: In vitro Assessment of Candidate Strains for SynCom Assembly
Purpose: To functionally characterize individual bacterial strains for the bottom-up construction of a stable, multifunctional SynCom [6].
Methodology:
Functional Phenotyping:
- Nitrogen Fixation Assay: Inoculate strains in nitrogen-free semi-solid medium. Quantify activity using the acetylene reduction assay, measuring ethylene production by gas chromatography. Express results in nmol C₂H₄ produced per hour per mg of protein [6].
- Phosphate Solubilization: Spot-inoculate strains onto National Botanical Research Institute's Phosphate (NBRIP) growth medium. Incubate and measure the solubilized phosphorus concentration in the medium (e.g., via the molybdenum-blue method), reporting results in mg/L [6].
- Indoleacetic Acid (IAA) Production: Grow strains in low-salt Luria-Bertani medium supplemented with 1 g/L tryptophan. After incubation, quantify IAA in the supernatant colorimetrically using Salkowski's reagent, reporting concentration in mg/L [6].
- Siderophore Production: Grow strains in Chrome Azurol S (CAS) agar plates. Measure the diameter of the orange halos formed, indicating siderophore production [6].
Metabolic Profiling:
- Utilize phenotype microarrays (e.g., Biolog plates) containing 58 carbon sources common to the target habitat (e.g., plant rhizosphere) [6].
- Inoculate each well with a standardized cell suspension and monitor colorimetric changes.
- Calculate the Resource Utilization Width (total number of carbon sources used) and the Resource Utilization Overlap (proportion of substrates shared between strains) [6].
Antagonistic Interaction Screening:
- Conduct cross-streak assays on solid media to evaluate interference competition.
- Visually inspect for zones of growth inhibition between strains to ensure negligible antagonistic effects prior to consortium assembly [6].
Protocol 2: Automated DBTL Cycle for Pathway Optimization
Purpose: To automate the DBTL cycle for high-throughput combinatorial optimization of genetic parts or pathways within a microbial host [32].
Methodology:
Design:
- Define a DNA library of components (e.g., promoters, Ribosome Binding Sites - RBS) for a pathway of interest.
- Use computational tools (e.g., UTR Designer) to generate a library of DNA sequences encoding different expression levels [33].
Build:
Test:
Learn:
- Collect and preprocess experimental data.
- Train machine learning models (e.g., Random Forest, Gradient Boosting) on the dataset to predict strain performance based on genetic design [31].
- Use an automated recommendation algorithm to select the most promising designs for the next DBTL cycle, balancing exploration and exploitation [31].
Quantitative Data and Analysis
The following tables summarize key quantitative metrics essential for analyzing and optimizing SynComs and metabolic pathways within the DBTL framework.
Table 1: Functional Phenotyping of Plant-Beneficial Bacterial Strains for SynCom Design
| Bacterial Strain | Nitrogen Fixation (nmol C₂H₄ h⁻¹ mg⁻¹) | Phosphate Solubilization (mg/L) | IAA Production (mg/L) | Siderophore Production |
|---|---|---|---|---|
| Azospirillum brasilense K | 3517 | Negligible | >40 | Low |
| Pseudomonas stutzeri G | 890 | 25.51 - 30.47 | 66.08 | High |
| Pseudomonas fluorescens J | Not detected | 46.39 | >40 | High |
| Bacillus velezensis SQR9 | Not detected | 25.51 - 30.47 | >40 | High |
| Bacillus megaterium L | Not detected | 25.51 - 30.47 | >40 | High |
| Cellulosimicrobium cellulans E | Not detected | Negligible | <40 | Low |
Data adapted from [6]
Table 2: Metabolic Interaction Metrics for SynCom Stability Analysis
| Strain Type | Example Strains | Avg. Resource Utilization Width | Avg. Metabolic Interaction Potential (MIP) | Avg. Metabolic Resource Overlap (MRO) |
|---|---|---|---|---|
| Narrow-Spectrum Resource (NSR) Utilizers | C. cellulans E, P. stutzeri G | 13.10 - 25.59 | 1.53 (High) | 0.51 (Low) |
| Broad-Spectrum Resource (BSR) Utilizers | B. velezensis SQR9, P. fluorescens J | 35.50 - 37.32 | 0.6 (Low) | 0.72 - 0.83 (High) |
Data synthesized from [6]. Note: NSR strains correlate with higher community stability.
Visualizations
DBTL Cycle Workflow for Community Optimization
Metabolic Interactions Governing Community Stability
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential Reagents and Materials for DBTL-based SynCom Research
| Item | Function/Application in DBTL Cycle | Specific Example / Note |
|---|---|---|
| Phenotype Microarrays | High-throughput profiling of carbon source utilization in the Design and Learn phases. | Biolog plates with 58 rhizosphere-relevant carbon sources to calculate Resource Utilization Width and Overlap [6]. |
| Genome-Scale Metabolic Models (GSMMs) | Computational prediction of metabolic interactions (MIP, MRO) during the Design phase. | Models refined with experimental phenotyping data; used to simulate all possible community combinations [6]. |
| Automated Strain Construction Platform | High-throughput Build phase for genetic manipulation and pathway optimization. | Laboratory robotics for DNA assembly, cloning, and transformation; enables combinatorial library construction [33] [32]. |
| Cell-Free Protein Synthesis (CFPS) System | In vitro Test phase for rapid prototyping of enzyme expression levels and pathway balance. | Crude cell lysate systems to bypass whole-cell constraints before in vivo testing [33]. |
| RBS Library Kit | Fine-tuning gene expression in metabolic pathways during the Build phase. | Library of Shine-Dalgarno sequences for modulating translation initiation rates without altering secondary structure [33]. |
| Machine Learning Algorithms | Data analysis and predictive model generation in the Learn phase. | Gradient Boosting and Random Forest models are robust for recommending designs in the low-data regime of early DBTL cycles [31]. |
A profound shift is occurring in microbial ecology, moving from simply cataloging which microorganisms are present to understanding what they are doing and how they interact. While traditional co-occurrence networks based on statistical correlations have provided valuable insights, they often fall short of revealing the underlying metabolic mechanisms governing interspecies interactions [34]. In the context of synthetic microbial communities (SynComs)—artificially created consortia of selected species—this mechanistic understanding is crucial for rational design aimed at improving stability and functionality [2] [35]. Genome-scale metabolic models (GEMs) have emerged as a powerful computational framework to address this challenge by simulating the complete metabolic network of microorganisms, enabling quantitative prediction of interaction outcomes [12] [36].
Flux Balance Analysis (FBA) stands as a cornerstone mathematical approach for analyzing GEMs. FBA computes the flow of metabolites through metabolic networks by optimizing an objective function (typically biomass production) under steady-state and mass-balance constraints [37]. This methodology has been extended to microbial communities through various frameworks that handle the complex trade-offs between individual species fitness and community-level fitness [37]. Among the specialized tools developed for community-level metabolic interaction analysis, SMETANA (Species MEtabolic Interaction ANalysis) offers a sophisticated algorithm for quantifying metabolic interactions by calculating the overlap and exchange of metabolic resources between community members [34].
These modeling approaches are particularly valuable for SynCom design, where predicting stable, multifunctional communities remains challenging. Metabolic modeling helps identify strains with complementary metabolic capabilities, potentially reducing competitive interactions while enhancing cooperative cross-feeding [6]. By integrating computational predictions with experimental validation, researchers are establishing a more rational framework for designing microbial consortia with predictable behaviors for agricultural, biomedical, and industrial applications [2].
Theoretical Foundations and Key Metrics
Fundamentals of Flux Balance Analysis for Communities
Flux Balance Analysis operates on the principle of stoichiometric mass balance, requiring that the production and consumption of each metabolite within a system are balanced at steady state. This is mathematically represented as S·v = 0, where S is the stoichiometric matrix containing stoichiometric coefficients of all reactions, and v is the flux vector representing reaction rates [37]. The solution space is constrained by lower and upper bounds on reaction fluxes (e.g., substrate uptake rates). FBA then identifies an optimal flux distribution that maximizes a cellular objective, most commonly biomass production.
When extended to microbial communities, FBA must account for metabolic interactions between species, primarily through metabolite exchange. The OptCom framework addresses this through a multi-level optimization formulation that explicitly considers trade-offs between individual species fitness and community-level fitness [37]. Unlike earlier approaches that relied on single objective functions, OptCom formulates separate biomass maximization problems for each species (inner problems) while optimizing a community-level objective function (outer problem). This structure enables OptCom to capture any combination of positive (mutualism, commensalism) and negative (competition) interactions within communities of any size [37].
Dynamic FBA (DFBA) further extends this approach by incorporating time-dependent changes in the extracellular environment [38]. DFBA formulates extracellular mass balances for key substrates and products and solves the coupled system of differential equations and linear programming problems, allowing researchers to predict population dynamics and metabolic shifts over time [38].
SMETANA: Algorithm and Quantitative Metrics
SMETANA implements a novel algorithm to quantify two key aspects of metabolic interactions in microbial communities: metabolic resource overlap and metabolic interaction potential [34]. The method goes beyond binary interaction predictions by providing continuous scores that reflect the strength and nature of metabolic interactions.
The SMETANA score quantifies the likelihood of metabolite exchange between community members. It calculates the proportion of metabolic secretions from one species that can be utilized by other community members, weighted by the importance of these metabolites for the recipient's metabolic network [34]. Mathematically, for a community with N species, the SMETANA score is computed as:
[ \text{SMETANA} = \frac{1}{N} \sum{i=1}^{N} \frac{1}{|Mi|} \sum{m \in Mi} \min\left(1, \sum{j \neq i} \delta{ijm}\right) ]
where (Mi) is the set of metabolites secreted by species *i*, and (\delta{ijm}) indicates whether metabolite m secreted by species i can be utilized by species j.
SMETANA also computes a Metabolic Resource Overlap (MRO) index, which quantifies competition for environmental resources by calculating the similarity in nutrient uptake profiles between community members [6]. Lower MRO values indicate reduced competition, while higher values suggest increased competitive pressure.
Table 1: Key Quantitative Metrics in Metabolic Interaction Analysis
| Metric | Calculation Approach | Interpretation | Application Context |
|---|---|---|---|
| SMETANA Score | Quantifies cross-feeding potential based on metabolite complementarity | Ranges 0-1; Higher values indicate stronger metabolic cooperation | Predicting cooperative interactions in SynCom design |
| Metabolic Resource Overlap (MRO) | Measures similarity in nutrient uptake profiles between strains | Higher values indicate increased competition for resources | Assessing competitive pressures in community assembly |
| Metabolic Interaction Potential (MIP) | Computes potential for cooperative metabolite exchange | Higher values suggest greater potential for cross-feeding | Identifying metabolically complementary strains |
| Metabolic Distance | Calculated using parsimonious Flux Balance Analysis (pFBA) | Quantifies metabolic similarity/differences between strains | Determining functional redundancy in communities |
Computational Protocols and Workflows
Integrated Analysis Pipeline: iNAP 2.0
The integrated Network Analysis Pipeline (iNAP 2.0) provides a user-friendly, web-based platform that incorporates SMETANA alongside other metabolic modeling tools for comprehensive interaction analysis [34]. This pipeline structures the analysis into four modular steps:
Module I: Prepare Genome-Scale Metabolic Models
- Input: Genome sequences (FASTA format) or annotated protein sequences (FAA format)
- Annotation: Prokka with default settings for gene annotation and identification of coding sequences
- Model Reconstruction: CarveMe for automated construction of genome-scale metabolic models from protein sequences
- Quality Control: Gap-filling function to correct models derived from metagenome-assembled genomes (MAGs) that may lack certain reactions due to binning or annotation limitations
- Output: SBML-FBC2 format models compatible with constraint-based modeling tools [34]
Module II: Infer Pairwise Interactions
- Method Selection: Choice of PhyloMint (phylogenetic distance-adjusted complementarity), SMETANA (cross-feeding prediction), or metabolic distance (pFBA-based)
- SMETANA Configuration: Set parameters for intracellular metabolites, exchange reactions, and nutritional constraints
- Computation: Distributed calculation of interaction metrics across all pairwise combinations
- Output: Numerical matrices of interaction indices (complementarity, competition, SMETANA scores) [34]
Module III: Construct Metabolic Interaction Networks
- Threshold Determination: Application of Random Matrix Theory (RMT) to identify statistically significant thresholds for network construction, moving beyond arbitrary cutoffs
- Network Assembly: Creation of microbial interaction networks where nodes represent species and edges represent significant metabolic interactions
- Metabolite Integration: Incorporation of potentially transferable metabolites as intermediate nodes to construct microbe-metabolite bipartite networks [34]
Module IV: Analyze Network Properties
- Topological Analysis: Calculation of standard network metrics (degree centrality, betweenness, clustering coefficient)
- Hub Identification: Determination of keystone species based on network connectivity
- Visualization: Generation of interactive network diagrams for exploratory analysis [34]
Figure 1: iNAP 2.0 Workflow for Metabolic Interaction Analysis
SMETANA Implementation Protocol
Step 1: Model Preparation and Curation
- Obtain genome-scale metabolic models for all community members in SBML format
- Ensure consistency in metabolite and reaction identifiers across models (recommended: BiGG database convention)
- Verify model quality through growth simulations on standard media
- For gap-filled models, document added reactions and nutritional constraints
Step 2: Define Nutritional Environment
- Specify available nutrients in the growth medium composition
- Set appropriate uptake bounds for each nutrient based on environmental conditions
- For root-associated communities, use root exudate-mimicking growth media containing typical plant-derived compounds [39]
Step 3: SMETANA Computation
- Run SMETANA algorithm to calculate metabolic interaction scores
- Compute both global (community-wide) and pairwise interaction potentials
- Generate metabolic resource overlap matrix for competitive interactions
Step 4: Result Interpretation
- Identify high-scoring metabolite exchanges that represent potential cross-feeding interactions
- Detect metabolic dead-ends where community functionality may be limited
- Pinpoint competitive bottlenecks where multiple species rely on the same limited resource
Application Case Studies
Agricultural SynCom Design for Plant Growth Promotion
A 2025 study demonstrated the power of metabolic modeling for constructing stable, multifunctional synthetic communities for agricultural applications [6]. Researchers selected six plant-beneficial bacterial strains with distinct functions (nitrogen fixation, phosphate solubilization, IAA synthesis, siderophore production) and analyzed their metabolic profiles using phenotype microarrays targeting 58 carbon sources commonly found in the plant rhizosphere.
Genome-scale metabolic models for each strain were refined using experimental phenotype data and applied to simulate all 57 possible community combinations (from two to six members each) [6]. SMETANA-based analysis revealed that strains with narrow-spectrum resource utilization (NSR) profiles, such as Cellulosimicrobium cellulans E and Pseudomonas stutzeri G, contributed significantly to elevated metabolic interaction potential (average MIP = 1.53), while broad-spectrum resource-utilizing (BSR) strains were associated with lower MIP scores (average = 0.6) [6].
Table 2: Metabolic Interaction Analysis of Plant-Beneficial Strains
| Bacterial Strain | Resource Utilization Width | Average MIP in Pairwise Communities | Key Functional Traits |
|---|---|---|---|
| Cellulosimicrobium cellulans E | 13.10 | 1.82 | IAA synthesis, metabolic specialization |
| Pseudomonas stutzeri G | 25.59 | 1.64 | Nitrogen fixation, IAA synthesis |
| Azospirillum brasilense K | 24.37 | 1.14 | Nitrogen fixation |
| Bacillus velezensis SQR9 | 35.50 | 0.55 | Phosphate solubilization, siderophore production |
| Bacillus megaterium L | 36.76 | 0.58 | Phosphate solubilization, IAA synthesis |
| Pseudomonas fluorescens J | 37.32 | 0.67 | Phosphate solubilization, siderophore production |
The resulting SynComs (SynCom4 and SynCom5) exhibited high stability in the tomato rhizosphere and increased plant dry weight by over 80%, demonstrating the practical value of metabolic modeling for designing effective agricultural inoculants [6].
Cheese Ripening Community Dynamics
Metabolic modeling has also proven valuable in food biotechnology applications. A 2024 study characterized the metabolism of a three-species community (Lactococcus lactis, Lactobacillus plantarum, and Propionibacterium freudenreichii) during a seven-week cheese production process [40]. Researchers used genome-scale metabolic models and omics data integration to model and calibrate individual dynamics using monoculture experiments, then coupled these models to capture community metabolism.
The dynamic model accurately predicted community dynamics and revealed the contribution of each microbial species to organoleptic compound production [40]. Metabolic exploration identified key interactions between bacterial species, including cross-feeding of metabolites that influence flavor development. This case study highlights how metabolic models can capture temporal dynamics in communities with industrial relevance.
Minimal Community Selection for Crop Plants
A 2023 study employed multi-genome metabolic modeling of 270 metagenome-assembled genomes from Campos rupestres to design a minimal synthetic microbial community to improve the yield of important crop plants [39]. Using the metage2metabo computational toolbox, researchers applied a targeted approach to select a minimal community encompassing essential compounds for microbial metabolism and compounds relevant to plant interactions.
This approach reduced the initial community size by approximately 4.5-fold while retaining crucial genes associated with essential plant growth-promoting traits, including iron acquisition, exopolysaccharide production, potassium solubilization, nitrogen fixation, GABA production, and IAA-related tryptophan metabolism [39]. The in-silico selection identified six hub species with notable taxonomic novelty that served as core components for stable SynComs.
Essential Research Toolkit
Table 3: Research Reagent Solutions for Metabolic Interaction Studies
| Tool/Resource | Function | Application Context |
|---|---|---|
| iNAP 2.0 | Web-based platform for metabolic interaction analysis | Integrated analysis of SMETANA, PhyloMint, and metabolic distance |
| CarveMe | Automated reconstruction of genome-scale metabolic models | Model building from protein sequences with gap-filling capability |
| ModelSEED | Alternative platform for metabolic model reconstruction | Creating models from genome annotations |
| Prokka | Rapid annotation of microbial genomes | Identification of coding sequences for downstream model construction |
| Cobrapy | Python library for constraint-based modeling | FBA simulation and analysis of metabolic models |
| Biolog Phenotype Microarrays | Experimental profiling of carbon source utilization | Validation and refinement of metabolic models |
| BiGG Database | Curated metabolic reaction database | Standardizing metabolite and reaction identifiers |
| PathwayTools | Pathway visualization and analysis | Metabolic network exploration and debugging |
Figure 2: Metabolic Modeling Methods and Their Primary Applications
SMETANA and Flux Balance Analysis provide powerful computational frameworks for quantifying metabolic interactions in synthetic microbial communities. By moving beyond statistical correlations to mechanistic, metabolism-based interaction predictions, these approaches enable more rational design of stable, functional SynComs. The integration of these computational methods with experimental validation through platforms like iNAP 2.0 represents a significant advancement in our ability to engineer microbial communities for agricultural, industrial, and biomedical applications.
As the field progresses, key challenges remain, including improving model accuracy through integration of multi-omics data, accounting for spatial organization in communities, and predicting long-term evolutionary dynamics [2]. Nevertheless, the continued refinement of metabolic modeling approaches promises to enhance our fundamental understanding of microbial interactions while providing practical tools for harnessing the power of microbial communities to address global sustainability challenges.
The rational design and analysis of synthetic microbial communities (SynComs) represent a frontier in microbial ecology and metabolic engineering. A significant challenge in this field lies in moving beyond descriptive studies to the predictive, model-driven manipulation of community structure and function. The integration of multi-omics data—specifically metagenomics, which defines community genetic potential and taxonomic composition, and proteogenomics, which links genomic information to expressed protein functions—is pivotal for closing this gap. This Application Note details protocols for the systematic integration of proteogenomic and metagenomic data into constraint-based metabolic models of SynComs. Framed within comparative metabolic modeling research, these methods enable researchers to generate mechanistic, testable hypotheses about community interactions, stability, and functional output, thereby accelerating the engineering of robust microbial consortia for biomedical and biotechnological applications.
Background
The Role of Multi-Omics in Decoding Microbial Communities
Microbial communities function as complex, integrated systems where metabolic capabilities emerge from the interactions between constituent members. While metagenomic sequencing characterizes the taxonomic composition and functional gene potential of a community, it provides limited insight into which pathways are actively operating in situ [41]. Proteogenomics, which couples genomic data with mass spectrometry-based proteomic profiling, directly identifies and quantifies the proteins expressed by a community, thereby reflecting its immediate functional state [42]. The integration of these complementary data layers with computational models transforms static genomic inventories into dynamic, predictive frameworks.
This integrated approach is particularly powerful for drug discovery and development, where understanding complex host-microbe and microbe-microbe interactions is essential. Network-based integration of multi-omics data can capture complex interactions between drugs and their multiple targets, improving predictions of drug responses, identifying novel drug targets, and facilitating drug repurposing [42]. Furthermore, the application of Model-Informed Drug Development (MIDD) frameworks, including Quantitative Systems Pharmacology (QSP), leverages such mechanistic models to inform quantitative decisions on drug dose, timing, and sequence [43] [44].
Metabolic Modeling of Synthetic Communities
Genome-scale metabolic models (GEMs) are mathematical representations of the metabolic network of an organism, enabling the simulation of metabolic fluxes under different conditions. For microbial communities, GEMs can be reconstructed and simulated using various approaches, each with distinct advantages [27]:
- Compartmentalized Models: Individual GEMs for each species are combined into a single stoichiometric matrix, with each species assigned a distinct compartment. This approach is most suitable for understanding organism-level interactions within a community.
- Mixed-Bag Approach: All metabolic pathways and transport reactions are integrated into a single model with one cytosolic and one extracellular compartment. This is best for analyzing interactions between different communities.
- Costless Secretion: Models are simulated using a dynamically updated medium based on exchange metabolites within the community.
A critical challenge has been the selection and reconciliation of automated tools for GEM reconstruction, as different tools rely on different biochemical databases. A comparative analysis revealed that tools like CarveMe, gapseq, and KBase, while using the same starting genomes, produce models with varying numbers of genes, reactions, and metabolic functionalities [27]. Consensus reconstruction methods, which combine the outcomes of multiple tools, have been shown to generate more comprehensive and functionally capable models, reducing bias and the presence of dead-end metabolites [27].
Key Reagents and Computational Tools
Table 1: Essential Research Reagents and Solutions for Omics Integration
| Category | Item/Software | Function/Description |
|---|---|---|
| Metagenomic Profiling | Meteor2 [41] | A tool for comprehensive Taxonomic, Functional, and Strain-level Profiling (TFSP) using environment-specific microbial gene catalogues. |
| Metabolic Reconstruction | CarveMe [27] | An automated tool for top-down reconstruction of GEMs from a universal template. |
| gapseq [27] | An automated tool for bottom-up reconstruction of GEMs, incorporating comprehensive biochemical data. | |
| KBase [27] | A platform offering bottom-up reconstruction of GEMs using the ModelSEED database. | |
| Model Reconciliation & Simulation | COMMIT [27] | A pipeline for gap-filling and refining draft community metabolic models. |
| COBRA Toolbox | A MATLAB suite for constraint-based reconstruction and analysis of metabolic models. | |
| Data Integration & Analysis | BioBakery Suite [41] | An all-in-one platform for TFSP, including MetaPhlAn (taxonomy) and HUMAnN (function). |
| KEGG Database [41] | A resource for functional orthology (KO) assignments and pathway mapping. | |
| dbCAN3 [41] | A tool for annotating carbohydrate-active enzymes (CAZymes). |
Integrated Experimental and Computational Protocol
This protocol outlines a complete workflow for building a proteogenomics-informed metabolic model of a synthetic microbial community.
Protocol 1: Community Profiling and Omics Data Acquisition
Objective: To generate high-quality metagenomic and metaproteomic data from a SynCom for model reconstruction and validation.
Materials:
- Synthetic Microbial Community sample
- DNA/Protein extraction kits (e.g., DNeasy PowerSoil Pro Kit, appropriate metaproteomic extraction buffers)
- Shotgun metagenomic sequencing platform (e.g., Illumina)
- LC-MS/MS system for metaproteomics
Procedure:
- Sample Harvesting: Culture the SynCom under defined experimental conditions. Harvest cells by centrifugation at a specified time point of interest (e.g., mid-log phase, stationary phase). Flash-freeze pellets in liquid nitrogen for omics analysis.
- DNA Extraction & Metagenomic Sequencing:
- Extract high-molecular-weight genomic DNA from the pellet.
- Prepare a shotgun metagenomic library following manufacturer protocols.
- Sequence using an Illumina platform to a minimum depth of 10 million paired-end reads (e.g., 2x150 bp) to ensure sufficient coverage for downstream analysis [41].
- Protein Extraction & Metaproteomic Analysis:
- From a parallel pellet, extract proteins using a suitable buffer.
- Digest proteins with trypsin and desalt the resulting peptides.
- Analyze peptides by LC-MS/MS on a high-resolution mass spectrometer.
- Search the resulting MS/MS spectra against a custom protein database generated from the SynCom's metagenomic assembly. This proteogenomic approach ensures accurate protein identification.
Protocol 2: Reconstruction of a Consensus Community Metabolic Model
Objective: To integrate outputs from multiple reconstruction tools into a unified, high-quality consensus GEM for the SynCom.
Materials:
- High-quality Metagenome-Assembled Genomes (MAGs) from the SynCom
- Computational tools: CarveMe, gapseq, KBase, and a consensus pipeline [27]
Procedure:
- Metagenomic Assembly and Binning: Assemble the quality-trimmed metagenomic reads into contigs. Bin contigs into MAGs using a tool like MetaBAT2. Refine MAGs to high-quality standards (completeness >90%, contamination <5%).
- Draft Model Reconstruction:
- Submit each high-quality MAG to the three automated reconstruction tools: CarveMe, gapseq, and KBase.
- Use default parameters for each tool. This will generate three distinct draft GEMs for each MAG.
- Consensus Model Generation:
- For each MAG, merge the three draft GEMs into a single draft consensus model using a reconciliation pipeline [27]. This process combines reactions and genes supported by multiple tools.
- The consensus model will typically encompass a larger number of reactions and metabolites while reducing dead-end metabolites compared to any single tool's output [27].
- Community Model Assembly:
- Combine the individual consensus GEMs for all MAGs into a compartmentalized community model. Each species model should occupy its own compartment, linked via a shared extracellular compartment.
- Gap-Filling with COMMIT:
- Perform gap-filling on the draft community model using the COMMIT tool [27].
- Initiate the process with a minimal medium definition. COMMIT will iteratively add transport reactions to enable the model to achieve growth by simulating the exchange of metabolites between community members.
Protocol 3: Model Refinement and Validation using Proteogenomic Data
Objective: To constrain and validate the consensus community model using quantitative metaproteomic data.
Materials:
- Consensus community metabolic model (from Protocol 2)
- Quantitative metaproteomic data (from Protocol 1)
- Constraint-based modeling software (e.g., COBRA Toolbox)
Procedure:
- Proteomic Data Mapping: Map identified proteins from the metaproteomic analysis to their corresponding enzyme-encoding genes in the consensus GEM.
- Model Constraining:
- Use the relative protein abundance levels as proxies for the maximum possible flux through the reactions catalyzed by those enzymes.
- Integrate these values as enzyme capacity constraints into the model. This step ensures that the model's flux predictions are biochemically realistic and reflect the community's actual proteomic state.
- Simulation and Validation:
- Simulate community growth or a target metabolic function (e.g., metabolite production) under the defined environmental conditions.
- Validate the model by comparing its predictions (e.g., secretion rates of specific acids, biomass formation) against experimentally measured data not used in the model construction.
- Iterative Refinement: Discrepancies between predictions and experimental data can guide targeted investigations into missing pathways, incorrect annotations, or poorly characterized interactions, initiating a new cycle of the Design-Build-Test-Learn (DBTL) framework.
The following diagram visualizes this integrated multi-protocol workflow.
Multi-Omics Model Integration Workflow
Data Analysis and Interpretation
Quantitative Analysis of Reconstruction Tools
The choice of reconstruction tool significantly impacts the structure and functional capacity of the resulting GEM. The following table summarizes a comparative analysis of models built from the same genomic input.
Table 2: Comparative Analysis of GEM Reconstruction Tools and Consensus Approach [27]
| Reconstruction Approach | Number of Reactions | Number of Metabolites | Number of Genes | Number of Dead-End Metabolites | Key Characteristics |
|---|---|---|---|---|---|
| CarveMe | Intermediate | Intermediate | Highest | Intermediate | Top-down approach; fast model generation using a universal template. |
| gapseq | Highest | Highest | Lowest | Highest | Bottom-up approach; comprehensive biochemical data integration. |
| KBase | Lowest | Lowest | Intermediate | Lowest | Bottom-up approach; uses ModelSEED database. |
| Consensus Model | High | High | High | Lowest | Integrates outputs from multiple tools; reduces bias and network gaps. |
Interpreting Proteogenomic Constraints
Integrating proteomic data transforms a generic metabolic network into a condition-specific model. The primary analysis involves using Flux Balance Analysis (FBA) to predict growth rates or metabolite exchange fluxes under proteomic constraints. Key outcomes include:
- Identification of Key Enzymes: Reactions whose fluxes are highly correlated with the abundance of their corresponding enzymes validate the model's mechanistic basis.
- Discovery of Metabolic Bottlenecks: Low flux through a pathway despite high enzyme abundance may indicate post-translational regulation or the need for cofactors not accounted for in the model.
- Elucidation of Interaction Dynamics: The model can predict cross-feeding metabolites. For example, it may show that one member's secretion of acetate is critical for another member's growth, a prediction that can be tested experimentally.
Table 3: Key Resources for SynCom Metabolic Modeling
| Resource Name | Type | Application in SynCom Research |
|---|---|---|
| Meteor2 [41] | Software | Provides integrated taxonomic, functional, and strain-level profiling from metagenomes, creating inputs for model reconstruction. |
| COMMIT [27] | Software/Pipeline | Performs gap-filling of community metabolic models to ensure metabolic functionality and network connectivity. |
| KEGG Modules [41] | Database/Annotation | Defines functional metabolic modules (e.g., Gut Metabolic Modules) used for functional profiling and model validation. |
| Genome-Scale Metabolic Model (GEM) | Conceptual Framework | The core mathematical representation of an organism's metabolism, serving as the building block for community models. |
| Design-Build-Test-Learn (DBTL) Cycle [2] | Engineering Framework | An iterative paradigm for the rational design and refinement of SynComs, where modeling drives hypothesis generation. |
| Ecological Interaction Principles [2] | Theoretical Foundation | Guides the selection of community members by engineering balanced cooperative and competitive relationships to enhance stability. |
Strategic Insights for Research Applications
The integration of proteogenomics and metagenomics into mechanistic models moves SynCom research from observational science to predictive engineering. The following diagram encapsulates the core ecological principles that should guide the initial design phase of a SynCom, which subsequently can be modeled and refined using the protocols described herein.
Ecological Design Principles for SynComs
Key strategic considerations for employing these protocols in a research program include:
- Embrace a Consensus Approach to Reconstruction: Relying on a single automated tool introduces database-specific biases. The consensus method demonstrably produces more comprehensive and functionally robust models, providing a superior foundation for simulation [27].
- Prioritize Model Validation: A model's predictive power is only as good as its validation. Always use orthogonal experimental data (e.g., product titers, nutrient consumption rates) not used in model building to test and refine model predictions.
- Leverage Models for Interaction Discovery: Use the proteogenomically-constrained model to generate hypotheses about critical metabolic interactions (e.g., which cross-fed metabolites are essential for community stability). These hypotheses can be tested by constructing simplified sub-communities or knock-out mutants.
- Adopt a DBTL Framework: Integrate this modeling workflow into an iterative DBTL cycle. Model predictions should inform the re-design of the SynCom (e.g., adding or removing species), leading to new rounds of testing, data generation, and model learning [2].
By adhering to these protocols and strategic principles, researchers can robustly integrate multi-omics data to construct predictive models of synthetic microbial communities, thereby accelerating the engineering of consortia with desired functions for therapeutic and industrial applications.
Synthetic Microbial Communities (SynComs) are precisely engineered consortia of microorganisms designed to mimic the functional attributes of natural microbiomes. The function-driven design paradigm represents a fundamental shift from taxonomy-based to function-based assembly, prioritizing the encoding and execution of key metabolic processes identified in target ecosystems [4]. This approach is foundational for comparative metabolic modeling research, as it enables the creation of tractable, hypothesis-driven model systems to dissect host-microbe and microbe-microbe interactions. By focusing on functional capacity over phylogenetic identity, researchers can construct SynComs that not only capture the ecological essence of complex microbiomes but also ensure cooperative coexistence and targeted functionality within specific host environments—from the human gut to the plant rhizosphere [4] [2]. This Application Note details the protocols and conceptual frameworks for designing, modeling, and experimentally validating host-tailored SynComs, providing a critical methodology for advancing synthetic ecology and microbiome engineering.
Core Principles and Workflow for Function-Driven SynCom Design
The function-driven design of SynComs is anchored in a multi-stage process that integrates computational prediction with experimental validation. The overarching goal is to select microbial strains that collectively encode a desired functional profile, derived from meta-omics data of a target habitat, and to ensure these strains can form a stable, interacting community within a specific host environment.
The following diagram illustrates the integrated workflow for designing a function-driven SynCom, from initial function identification to final experimental validation.
Key Computational and Experimental Stages
Stage 1: Functional Profiling: The process begins with a comparative analysis of metagenomic samples from the host environment of interest (e.g., healthy vs. diseased state). Proteins are predicted from metagenomic assemblies and annotated against functional databases (e.g., Pfam) [4]. Core functions (prevalent in >50% of samples) and differentially enriched functions (e.g., in diseased hosts) are identified and assigned weights to prioritize them during strain selection [4].
Stage 2: Strain Selection: The weighted functional profile is used to select an optimal set of strains from a comprehensive genome collection (e.g., isolate genomes or Metagenome-Assembled Genomes). Tools like the MiMiC2 algorithm score each genome based on its encoded Pfams, favoring those that match the metagenome's functional signature while minimizing redundant or extraneous functions [4].
Stage 3: In Silico Validation: Before experimental assembly, the proposed community is modeled in silico. Genome-scale metabolic models (GEMs) of each member are constructed and simulated using platforms like BacArena or Virtual Colon [4] [12]. This step predicts cooperative growth, metabolic interactions (e.g., cross-feeding), and overall community stability, providing critical evidence for coexistence [4] [28].
Stage 4: Experimental Validation: The final, critical stage involves physically constructing the SynCom and testing its function and stability in a relevant host model, such as gnotobiotic mice or axenic plants [4] [6]. The community's impact on host phenotype (e.g., induction of colitis or plant growth promotion) is assessed to confirm its functional efficacy [4] [6].
Quantitative Metrics for Community Design and Stability
Metabolic modeling provides quantitative metrics to guide the rational design of stable SynComs. Two key indices, Metabolic Interaction Potential (MIP) and Metabolic Resource Overlap (MRO), are critical for predicting community dynamics from genomic data.
Table 1: Key Quantitative Metrics for SynCom Design and Evaluation
| Metric | Description | Computational Tool | Interpretation and Impact on Stability |
|---|---|---|---|
| Metabolic Interaction Potential (MIP) | Quantifies the potential for cooperative cross-feeding and metabolic interdependence between community members [6]. | Genome-scale metabolic models (GEMs), SMETANA [28] [6] | Higher MIP indicates stronger potential for cooperation, enhancing community stability and function [6]. |
| Metabolic Resource Overlap (MRO) | Measures the degree of similarity in resource utilization profiles among member strains, indicating niche overlap [6]. | GEMs, constrained by phenotypic data (e.g., Biolog arrays) [6] | Lower MRO reduces direct competition for nutrients, thereby increasing the likelihood of stable coexistence [6]. |
| Resource Utilization Width | Reflects the diversity of carbon or nitrogen sources a strain can use [6]. | Phenotype microarrays (e.g., Biolog) [6] | Narrow-spectrum utilizes show higher MIP and lower MRO, correlating with greater stability [6]. |
The relationship between a strain's metabolic niche and its role in the community is a key design consideration. Studies show that narrow-spectrum resource-utilizing (NSR) strains—those with specialized metabolic capabilities—often serve as central nodes in the community's metabolic network. For instance, Cellulosimicrobium cellulans and Pseudomonas stutzeri, both NSR strains, were found to enhance community stability by secreting key metabolites like asparagine, vitamin B12, and isoleucine, thereby fostering metabolic interdependence [6]. In contrast, broad-spectrum resource-utilizing (BSR) strains tend to have higher MRO, leading to increased competitive pressure within the consortium [6].
Detailed Experimental Protocols
Protocol 1: High-Throughput Construction of SynCom Variants
Manually assembling all possible combinations of strains from a candidate pool is necessary for comprehensively testing community assembly rules. This protocol enables the systematic construction of hundreds to thousands of unique SynComs in a microtiter plate format [45].
Principle: The protocol leverages combinatorial mathematics to assign each unique SynCom combination a specific well location on a microtiter plate. The total number of combinations for n strains is 2n, which includes all subsets from single species to the full consortium, plus a blank control [45].
Materials and Reagents:
- Candidate Bacterial Strains: 4-11 distinct, cultured strains [45].
- Microtiter Plates: 24-well, 96-well, or 384-well plates, depending on the scale [45].
- Liquid Culture Media: Appropriate for all strains (e.g., TSB, LB) [45].
- Pipettes: Single-channel and multi-channel (8 or 16-channel) pipettes [45].
- Reagent Reservoirs [45].
- Software: The
synconsR package for generating plate maps and unique SynCom IDs [45].
Procedure:
- Strain Preparation: Grow each candidate strain in liquid medium to a standardized optical density (e.g., OD600 = 1.0).
- Experimental Design: Run the
synconsR package to generate a detailed plate map. The output will assign a unique ID to each well and specify which strains need to be added to that well to create a specific SynCom [45]. - Sequential Inoculation:
- Arrange the cultures of the candidate strains in a specific, fixed order (e.g., S1, S2, S3... S11).
- Using a multi-channel pipette, sequentially add each strain to all wells designated to contain it according to the plate map. This systematic approach minimizes pipetting errors and cross-contamination [45].
- After all additions, top up each well with sterile medium to the final required volume.
- Incubation and Tracking: Incubate the plates under appropriate conditions. Use the generated data collection forms from the
synconspackage to track the composition and subsequent experimental results for each well [45].
Protocol 2: In Silico Community Simulation with BacArena
Metabolic modeling with BacArena provides a cost-effective method to simulate community dynamics and predict stability prior to resource-intensive experimental work [4].
Principle: BacArena is a computational tool that integrates genome-scale metabolic models (GEMs) into a spatial, agent-based simulation framework. It models individual bacterial cells and their metabolic exchanges within a shared environment, predicting growth and interaction dynamics over time [4].
Materials and Software:
Procedure:
- Model Reconstruction: Generate a high-quality GEM for each SynCom member from its genome sequence using GapSeq (command:
doall) [4]. - Arena Setup: In R, create an Arena object (e.g., size 100x100) using the
Arena()command. Add a default, non-specific medium to the arena usingaddDefaultMed()to simulate a generic, permissive environment [4]. - Inoculation: Depending on the script used:
- SingleGrowth.R: Add 10 cells of a single member to the arena (
addOrg()). - PairedGrowth.R: Add 10 cells each of two members to study pairwise interactions.
- Combined_Growth.R: Add 10 cells of all SynCom members to simulate the full community [4].
- SingleGrowth.R: Add 10 cells of a single member to the arena (
- Simulation Execution: Run the simulation for a defined period (e.g., 7 hours) using the
simEnv()command [4]. - Data Analysis: Extract growth data and metabolite exchange fluxes from the simulation output. Analyze these results to infer cooperation, competition, and the potential for stable coexistence.
- Model Reconstruction: Generate a high-quality GEM for each SynCom member from its genome sequence using GapSeq (command:
Table 2: Key Research Reagents and Computational Tools for SynCom Development
| Item Name | Type | Key Function in SynCom Research |
|---|---|---|
| MiMiC2 Algorithm | Software/Bioinformatics Pipeline | Automated, function-based selection of SynCom members from genome collections using weighted metagenomic functional profiles [4]. |
| BacArena Toolkit | Software/Metabolic Modeling Platform | Agent-based simulation of multi-species community dynamics by integrating GEMs, predicting growth and interactions in silico [4]. |
| GapSeq | Software/Metabolic Modeling Tool | Automated reconstruction of high-quality genome-scale metabolic models (GEMs) from genomic data, serving as input for BacArena [4]. |
syncons R Package |
Software/Experimental Design Tool | Generates unique IDs and plate maps for the high-throughput, manual construction of thousands of SynCom combinations in microplates [45]. |
| Phenotype Microarrays (e.g., Biolog) | Laboratory Assay | High-throughput profiling of strain resource utilization (e.g., 58 carbon sources) to calculate Resource Utilization Width and Overlap [6]. |
| Virtual Colon Toolkit | Software/Host-Microbe Modeling | A specialized modeling environment for simulating microbial community dynamics within the physiologically structured environment of the human colon [4]. |
The function-driven design of SynComs, powered by comparative metabolic modeling, provides a robust and predictive framework for engineering host-associated microbial communities. By prioritizing functional capacity, quantitatively assessing metabolic interactions, and employing high-throughput experimental validation, researchers can move beyond descriptive ecology to predictive ecosystem engineering. The protocols and tools outlined in this Application Note provide a concrete path for constructing SynComs that are not only taxonomically defined but also functionally representative and ecologically stable within their target host environment. This methodology is pivotal for advancing therapeutic, agricultural, and environmental applications of synthetic ecology.
Navigating Complexity: Overcoming Challenges in Model Accuracy and Community Stability
In the field of comparative metabolic modeling of synthetic microbial communities, genome-scale metabolic models (GEMs) serve as powerful computational frameworks to predict phenotypic outcomes from genotypic information and to understand metabolic interactions between organisms [13] [36]. The construction of high-quality GEMs is a critical first step, and several automated reconstruction tools have been developed to streamline this process. Among the most prominent are CarveMe, gapseq, and the KBase platform, each employing distinct algorithms, biochemical databases, and reconstruction philosophies [13] [46].
However, the choice of reconstruction tool is not neutral. Evidence indicates that different tools applied to the same genome can produce models with varying structural and functional properties, introducing a tool-based bias that can significantly impact downstream predictions about community metabolic capabilities and interactions [13]. This application note delineates the sources of bias among these three tools, provides a quantitative comparison of their outputs, and outlines experimental protocols for conducting a robust comparative analysis, thereby empowering researchers to make informed choices in their microbial community research.
Fundamental Reconstruction Philosophies and Databases
The core differences between CarveMe, gapseq, and KBase stem from their foundational approaches to model building and the biochemical databases they utilize.
- CarveMe employs a top-down reconstruction strategy. It starts with a universal, curated metabolic network (a "template") and carves away reactions that are not supported by genomic evidence from the target organism [13]. This approach prioritizes network functionality and ensures a high degree of connectivity from the outset.
- gapseq utilizes a bottom-up approach. It constructs models from scratch by mapping annotated genomic sequences to a manually curated reaction database [13] [46]. A key feature is its novel gap-filling algorithm, which leverages both network topology and sequence homology to reference proteins to resolve metabolic gaps, aiming to reduce medium-specific effects during reconstruction [46].
- KBase (which implements the ModelSEED pipeline) is also a bottom-up tool that builds draft models from genome annotations [13] [47]. It is integrated into a comprehensive web-based platform that supports various -omics data analyses and modeling workflows [48].
The biochemical database underlying each tool is a primary source of variation. gapseq uses a dedicated, curated database derived from ModelSEED but extended and refined [46]. Both CarveMe and KBase rely on their own distinct databases (BiGG and ModelSEED, respectively), which use different namespaces for metabolites and reactions, creating challenges when combining models from different tools [13].
Quantitative Comparison of Model Properties
A comparative analysis of GEMs reconstructed from the same set of 105 marine bacterial metagenome-assembled genomes (MAGs) revealed significant structural differences [13]. The table below summarizes the key findings.
Table 1: Structural characteristics of community metabolic models built from the same MAGs using different reconstruction tools. Data adapted from [13].
| Reconstruction Tool | Reconstruction Philosophy | Number of Genes (Relative) | Number of Reactions & Metabolites | Number of Dead-End Metabolites | Similarity to Consensus Models (Jaccard Index for Genes) |
|---|---|---|---|---|---|
| CarveMe | Top-down | Highest | Intermediate | Lower | High (0.75-0.77) |
| gapseq | Bottom-up | Lowest | Highest | Higher | Information Not Specified |
| KBase | Bottom-up | Intermediate | Lower | Intermediate | Information Not Specified |
Further analysis of the similarity between tools showed that models generated by gapseq and KBase, which share a common database ancestry (ModelSEED), exhibited higher similarity in reaction and metabolite sets (Jaccard similarity of ~0.24 and ~0.37, respectively) compared to CarveMe models [13]. In contrast, CarveMe and KBase models showed greater similarity in their gene sets (Jaccard similarity of ~0.42-0.45) [13].
Performance in Phenotypic Predictions
The ultimate test of a metabolic model is its accuracy in predicting biological phenotypes. Independent benchmarking studies have evaluated these tools on various tasks:
- Enzyme Activity Prediction: When tested against a large dataset of 10,538 experimental enzyme activities from the Bacterial Diversity Metadatabase (BacDive), gapseq demonstrated a lower false negative rate (6%) and higher true positive rate (53%) compared to CarveMe (32% false negative, 27% true positive) and ModelSEED/KBase (28% false negative, 30% true positive) [46].
- Strain-Specific Growth Predictions: In a study focused on Klebsiella pneumoniae, a model generated by Bactabolize (a newer, reference-based tool) performed comparably or better than models from CarveMe and gapseq across 507 substrate utilization predictions and 2,317 knockout mutant growth predictions [48]. This highlights that the choice of tool can be organism and context-dependent.
- Utility in Community Modeling: A significant finding is that the set of exchanged metabolites predicted in a community context is more influenced by the reconstruction tool itself than by the specific bacterial community being studied [13]. This suggests a strong potential for bias when predicting metabolite cross-feeding using community GEMs.
Protocols for Comparative Tool Assessment
To systematically evaluate and mitigate tool-based bias in your research, follow this two-stage experimental protocol.
Protocol 1: Cross-Tool Model Reconstruction and Structural Analysis
Objective: To generate and structurally compare metabolic models for a target organism or community using CarveMe, gapseq, and KBase. Materials: A high-quality genome sequence (FASTA format) for your organism of interest. Software: CarveMe, gapseq, and an account on the KBase platform.
Procedure:
- Model Reconstruction:
- CarveMe: Run the
carvecommand on your genome file. Use the--initoption to specify a medium if needed. - gapseq: Execute the
gapseqpipeline withgapseq findandgapseq draftcommands. Thegapseq transcommand can be added to predict transport reactions. - KBase: Use the "Build Metabolic Model" app on the KBase platform, providing your genome as input.
- CarveMe: Run the
- Model Curation: Standardize the output models to a common notation system (e.g., MetaNetX) to enable direct comparison, as different tools use different metabolite and reaction identifiers [13].
- Structural Analysis: For each model, calculate and compile the following metrics into a table similar to Table 1 above:
- Total number of genes, reactions, and metabolites.
- Number of dead-end metabolites.
- Number of transport reactions.
- Core metabolic functionality (e.g., ability to produce essential biomass precursors).
Protocol 2: Functional Validation and Consensus Modeling
Objective: To assess the predictive performance of each model and leverage a consensus approach to mitigate individual tool bias. Materials: Experimentally determined phenotypic data for your organism(s), such as carbon source utilization or gene essentiality data.
Procedure:
- Phenotype Prediction: For each model, simulate growth phenotypes.
- Perform in silico Flux Balance Analysis (FBA) to predict growth on a panel of carbon sources.
- Simulate single-gene knockout mutants and predict essential genes.
- Validation: Compare the predictions from each tool against the experimental data to calculate accuracy, precision, recall, and F1-score.
- Consensus Model Generation:
- Use a pipeline like COMMIT to merge draft models originating from the same genome but built with different tools [13].
- Perform gap-filling on this merged draft community model to ensure functionality.
- Analysis: Evaluate the consensus model against the individual tool-generated models. The consensus model should encompass a larger number of reactions while reducing dead-end metabolites, leading to enhanced functional capability [13].
The following diagram visualizes this integrated workflow for assessing and mitigating tool-based bias.
Table 2: Key software and data resources for metabolic model reconstruction and analysis.
| Resource Name | Type | Function & Application |
|---|---|---|
| CarveMe [13] | Software Tool | Automated, top-down reconstruction of GEMs from a universal template. Prioritizes speed and network connectivity. |
| gapseq [13] [46] | Software Tool | Automated, bottom-up reconstruction with informed pathway prediction and gap-filling. Emphasizes biochemical database curation. |
| KBase [13] | Web Platform | Integrated environment for reconstruction (via ModelSEED) and analysis of metabolic models and microbial communities. |
| COBRApy [48] [49] | Software Library | Python toolbox for constraint-based modeling and simulation, used as the backend by many tools including Bactabolize. |
| COMMIT [13] | Software Tool | A pipeline for gap-filling and refining community metabolic models, useful for building consensus models. |
| MEMOTE [48] | Software Tool | A tool for assessing and ensuring the quality of genome-scale metabolic reconstructions. |
| AGORA2 [47] | Model Resource | A curated resource of 7,302 manually refined microbial metabolic models, serving as a gold standard for the human gut microbiome. |
| BacDive [46] | Data Resource | A database containing experimental phenotypic data (e.g., enzyme activity, substrate use) for bacterial strains, used for model validation. |
The automated reconstruction tools CarveMe, gapseq, and KBase each present distinct strengths and biases arising from their underlying philosophies, databases, and algorithms. gapseq often shows superior accuracy in predicting enzyme activities and carbon source utilization, while CarveMe offers speed and high connectivity. KBase provides an integrated, user-friendly platform. Critically, the choice of tool can predetermine the predicted metabolic interactions in a community.
To achieve robust and reliable results in synthetic microbial community research, we recommend a consensus-driven approach. By systematically comparing models from multiple tools, validating predictions against experimental data where possible, and leveraging consensus-building techniques, researchers can effectively mitigate tool-based bias and unlock the full potential of metabolic modeling to understand and engineer microbial ecosystems.
The Consensus Model Approach to Reduce Uncertainty and Minimize Dead-End Metabolites
Genome-scale metabolic models (GEMs) are powerful mathematical representations of microbial metabolism that enable the prediction of cellular phenotypes from genomic information. However, the reconstruction of GEMs using different automated tools often results in models with significant structural and functional variations, introducing substantial uncertainty in model predictions and limiting their biological relevance [27]. This uncertainty stems from each reconstruction pipeline relying on distinct biochemical databases, annotation methods, and network building algorithms [50]. A promising solution to this challenge is the consensus model approach, which integrates multiple individual reconstructions into a unified model that captures the strengths of each method while mitigating their individual weaknesses [27]. This approach is particularly valuable for modeling synthetic microbial communities, where predictive accuracy is crucial for designing communities with desired metabolic functions [51].
The consensus approach directly addresses the pervasive problem of dead-end metabolites—metabolic compounds that cannot be produced or consumed by the network due to gaps in our biochemical knowledge. By combining evidence from multiple reconstruction sources, consensus models significantly reduce the number of these metabolically inaccessible compounds, leading to more complete and functional network representations [27]. This technical note details the methodology for constructing and validating consensus metabolic models, with specific applications for synthetic microbial community engineering.
Quantitative Advantages of Consensus Models
Comparative analyses of metabolic models reconstructed from the same genomes using different automated tools reveal striking structural differences that directly impact model functionality. The table below summarizes key quantitative improvements achieved through the consensus modeling approach:
Table 1: Structural Improvements in Consensus Metabolic Models
| Model Characteristic | Individual Models | Consensus Model | Improvement |
|---|---|---|---|
| Reaction Coverage | Variable between tools (gapseq > CarveMe > KBase) | Highest number of reactions | Increased comprehensiveness |
| Metabolite Inclusion | Variable between tools | Largest metabolite set | Enhanced network connectivity |
| Dead-end Metabolites | Highest in gapseq models | Significantly reduced | Improved network functionality |
| Genomic Evidence | Varies by tool (CarveMe has most genes) | Strongest genomic support | Increased biological relevance |
| Gene-Reaction Associations | Tool-dependent | Most comprehensive | Improved annotation integration |
Studies examining models from CarveMe, gapseq, and KBase revealed that despite being reconstructed from the same metagenome-assembled genomes (MAGs), these approaches yielded GEMs with distinct reaction sets, varying metabolite numbers, and different metabolic functionalities [27]. The Jaccard similarity for reaction sets between individual reconstructions was remarkably low (0.23-0.24 on average), highlighting the significant discrepancies between tools [27]. Consensus models address these limitations by encompassing a larger number of reactions and metabolites while concurrently reducing the presence of dead-end metabolites [27].
Protocol for Consensus Model Reconstruction
The following diagram illustrates the comprehensive workflow for constructing consensus metabolic models:
Detailed Experimental Procedures
Step 1: Multi-Tool Model Reconstruction
Begin by generating draft metabolic reconstructions using at least three different automated reconstruction tools. The selected tools should represent both top-down and bottom-up reconstruction philosophies:
- CarveMe: Uses a top-down approach, starting with a universal model and removing reactions without genomic evidence [27] [50]
- gapseq: Employs a bottom-up approach, mapping annotated genes to reactions from comprehensive biochemical databases [27] [50]
- KBase: Provides an integrated platform for reconstruction and analysis using the ModelSEED database [27]
- RAVEN 2.0: Leverages the KEGG and MetaCyc databases for reaction mapping [52]
Protocol Notes: Use identical genomic input data across all tools. For microbial communities, ensure consistent quality criteria for metagenome-assembled genomes (MAGs).
Step 2: Namespace Unification and Identifier Mapping
Convert all model components (metabolites, reactions, genes) to a common namespace to enable cross-tool comparison:
- Metabolite ID Conversion: Use MetaNetX or a similar platform to map metabolite identifiers across different databases (BiGG, ModelSEED, MetaCyc, KEGG) [50] [52]
- Reaction Standardization: Align reaction equations, directionality, and protonation states
- Gene Identifier Mapping: Use BLAST or similar sequence alignment tools to reconcile gene identifiers across annotations [50]
Critical Considerations: Track conversion success rates. Features that cannot be converted (stored in a "not_converted" field) may require manual curation [50].
Step 3: Consensus Generation
Integrate the standardized models using one of these computational approaches:
- GEMsembler Method: Employ the GEMsembler Python package to systematically combine models and generate consensus models with features present in a specified number of input models (e.g., core2 for features in ≥2 models) [50]
- Feature Confidence Assessment: Assign confidence levels to metabolites, reactions, and genes based on their presence across input models
- Attribute Agreement: Resolve conflicting reaction attributes (e.g., directionality) based on majority agreement among input models [50]
Step 4: Community-Specific Gap Filling
Apply the COMMIT algorithm to fill metabolic gaps while considering community composition and metabolite leakage:
- Metabolite Permeability: Prioritize secretion of metabolites based on membrane permeability predictions [52]
- Iterative Gap-Filling: Start with a minimal medium and dynamically update available metabolites based on community secretion profiles [27] [52]
- Order Independence: Note that the iterative order of gap-filling shows negligible correlation with the number of added reactions (r = 0-0.3), supporting methodological robustness [27]
Table 2: Key Resources for Consensus Model Development
| Resource Name | Type | Primary Function | Application Context |
|---|---|---|---|
| GEMsembler | Python Package | Cross-tool GEM comparison and consensus building | Structural analysis, model integration, and curation [50] |
| COMMIT | Algorithm | Community-aware gap filling | Considering metabolite leakage and permeability in communities [52] |
| MetaNetX | Platform | Database namespace reconciliation | Metabolite and reaction identifier mapping across sources [50] [52] |
| CarveMe | Reconstruction Tool | Top-down model reconstruction | Fast draft generation using universal templates [27] [50] |
| gapseq | Reconstruction Tool | Bottom-up model reconstruction | Comprehensive biochemical network mapping [27] [50] |
| KBase | Platform | Integrated reconstruction and analysis | User-friendly model building using ModelSEED [27] |
| CHESHIRE | Deep Learning Tool | Topology-based gap filling | Predicting missing reactions from network structure [53] |
Application in Synthetic Microbial Community Engineering
The consensus model approach provides particular value for designing and optimizing synthetic microbial communities (SynComs). The methodology enables more accurate prediction of metabolic interactions that drive community stability and function [51]. When engineering communities for specific biotechnological applications—such as production of high-value compounds, biodegradation of pollutants, or therapeutic interventions—consensus models of individual members reduce uncertainty in predicting cross-feeding relationships and resource competition [51] [54].
For human health applications, specifically developing live biotherapeutic products (LBPs), consensus models of gut microbes can guide the design of synthetic communities with defined metabolic capabilities [54]. These models help identify synergistic interactions that enhance community persistence and function in the gastrointestinal environment. Similarly, in agricultural and environmental applications, consensus models of plant-associated microbes support the design of communities that promote plant growth and stress resistance through more predictable metabolic interactions [55].
The reduced incidence of dead-end metabolites in consensus models is particularly crucial for community modeling, as these gaps can block the simulation of metabolic exchanges that sustain community members. By providing more complete network representations, consensus models enable more reliable identification of potential helpers (organisms that leak essential metabolites) and beneficiaries (organisms that consume these metabolites) within synthetic communities [52].
Validation and Quality Assessment
Structural Validation Metrics
Implement quantitative measures to assess consensus model quality:
- Jaccard Similarity Analysis: Compare sets of reactions, metabolites, and genes between individual and consensus models [27]
- Dead-end Metabolite Reduction: Quantify the decrease in blocked metabolites compared to individual reconstructions
- Genomic Support Evaluation: Verify that reactions in the consensus model maintain strong gene-protein-reaction (GPR) associations
Functional Validation
Assess model performance using biological relevant tests:
- Auxotrophy Prediction Accuracy: Compare model predictions of nutrient requirements with experimental data [50]
- Gene Essentiality Predictions: Evaluate how well consensus models predict essential genes compared to gold-standard curated models [50]
- Metabolic Interaction Validation: For community models, compare predicted metabolic exchanges with experimental measurements of metabolite secretion and consumption
Technical Considerations and Limitations
While the consensus approach offers significant advantages, several technical challenges require attention:
- Computational Complexity: Integrating multiple models and performing community-scale gap-filling demands substantial computational resources, particularly for large communities [52]
- Namespace Reconciliation: Despite tools like MetaNetX, incomplete mapping between biochemical databases can result in information loss during conversion [50]
- Conflict Resolution: Discrepancies in reaction directionality, GPR rules, and biomass composition between input models require systematic resolution protocols [50]
- Tool Selection Bias: The choice of reconstruction tools included in the consensus affects the final model; including tools with complementary strengths (e.g., CarveMe for speed, gapseq for comprehensiveness) is recommended [27]
Future methodology developments should focus on standardized conflict resolution protocols, improved database integration, and machine learning approaches to further enhance consensus model quality and predictive power [53].
Synthetic microbial communities (SynComs) are engineered multispecies systems that perform complex functions through division of labor, offering superior metabolic flexibility and functional stability compared to single-strain cultures [56]. However, their stability is perpetually threatened by two primary challenges: the emergence of social "cheaters" that exploit community resources without contributing to collective fitness, and uncontrolled competition that can drive functional collapse [57] [58]. This Application Note provides experimental methodologies and conceptual frameworks grounded in comparative metabolic modeling to address these stability challenges, enabling robust SynCom design for biomedical and biotechnological applications.
Theoretical Framework: Stability Mechanisms
Cheater Dynamics and Control Mechanisms
Social cheating occurs when antibiotic-sensitive strains benefit from public goods produced by resistant strains without bearing the metabolic cost of resistance mechanisms. In kin bacterial communities, this manifests as resistant "cooperator" strains detoxifying the environment for sensitive "cheater" strains, creating stability challenges when significant growth rate differences exist between strains [57]. Theoretical modeling and experimental validation with Comamonas testosteroni strains KF-1 (cooperator) and CNB-2/ΔLuxR (cheater) under sulfamethoxazole stress demonstrate that coexistence becomes possible only through carefully regulated interspecific interactions [57].
Interspecific Competition as a Stabilizing Force
Introducing a third species as a "regulator" can transform community dynamics from competitive exclusion to stable coexistence. This occurs through competitive interference rather than facilitation, where the external competitor mitigates intraspecific inhibition by redirecting competitive pressures [57]. In practice, Pseudomonas aeruginosa introduction into the C. testosteroni system created sufficient interspecific competition to balance the cooperator-cheater dynamics, enabling prolonged coexistence despite inherent growth rate disparities [57].
Table 1: Stabilization Mechanisms for Synthetic Microbial Communities
| Mechanism | Principle | Experimental Validation | Effect on Stability |
|---|---|---|---|
| Third-Party Competitor | Introduces interspecific competition to balance intraspecific dynamics | P. aeruginosa in C. testosteroni system [57] | Prevents competitive exclusion of cooperators by cheaters |
| Spatial Structuring | Creates physical niches that protect cooperators from exploitation | Bacillus subtilis starch digestion biofilms [58] | Enables cooperator refuge formation and local positive feedback |
| Metabolic Cross-Feeding | Establfficient mutual dependencies through metabolite exchange | Engineered auxotrophic S. cerevisiae strains [58] | Creates evolutionary constraints against pure cheating strategies |
| Quorum Sensing Regulation | Links public good production to population density | AHL-mediated denitrification control in wastewater SynComs [21] | Prevents wasteful production at low densities while ensuring sufficient production at high densities |
| Division of Labor | Distributess metabolic burden across specialized strains | Aerobic denitrification consortia under DBP/LOFX stress [21] | Enhances functional redundancy and resilience to perturbations |
Experimental Protocol: Full Factorial Community Assembly
Combinatorial Construction Workflow
This protocol enables systematic assembly of all possible strain combinations from a microbial library to identify optimal community compositions that maximize function while suppressing cheaters [59].
Materials and Equipment:
- Microbial strain library (pure cultures)
- Sterile 96-well plates
- Multichannel pipettes (10-100 μL, 100-1000 μL)
- Sterile nutrient broth medium
- Mineral salt medium (for experimental assays)
- Plate reader or flow cytometer for high-throughput biomass quantification
Procedure:
- Strain Preparation: Grow each library strain to mid-log phase in separate culture vessels. Adjust cell densities to standardized OD₆₀₀ = 0.5 for uniform inoculation.
- Binary Encoding: Assign each of m species a unique binary identifier (e.g., Species 1 = 00000001, Species 2 = 00000010) [59].
- Initial Plate Setup: In Column 1 of a 96-well plate, assemble all combinations of the first three species following binary numbering (000, 001, 010, 011, 100, 101, 110, 111).
- Iterative Expansion: Duplicate the contents of Column 1 to Column 2. Add Species 4 to all wells in Column 2 using a multichannel pipette.
- Progressive Assembly: Repeat duplication and addition process for remaining species, doubling the number of columns with each new species addition.
- Functional Screening: Incubate plates under appropriate conditions and measure target functions (biomass production, metabolite production, etc.).
- Interaction Mapping: Quantify all pairwise and higher-order interactions from functional data to identify optimal strain combinations.
Figure 1: Full Factorial Community Assembly Workflow. This systematic approach enables empirical mapping of community-function landscapes to identify optimal strain combinations that resist cheater invasion [59].
Application Note: Maintaining Functional Stability Under Disturbance
Case Study: Aerobic Denitrification Under Antibiotic Stress
A synthetic microbial community comprising Pseudomonas aeruginosa N2, Acinetobacter baumannii N1, and Aeromonas hydrophila demonstrated remarkable functional stability maintaining ~93% denitrification efficiency under dibutyl phthalate (DBP) and levofloxacin (LOFX) disturbances [21].
Key Stability Mechanisms Identified:
- Metabolic Network Rewiring: DBP and LOFX triggered distinct metabolic reprogramming, accelerating the TCA cycle to enhance energy flux and extracellular polymeric substance production.
- Electron Flux Redirection: DBP stimulated phenazine-1-carboxylic acid production, accelerating electron transfer from quinone pool to complex III.
- Quorum Sensing Mediation: Interspecific communication through C4-HSL and 3OC12-HSL facilitated labor division and coordination under stress.
- Functional Redundancy: Different species assumed functional dominance under different disturbances - AH and PA under DBP, AC and PA under LOFX - maintaining overall community function despite compositional shifts.
Table 2: Quantitative Stability Performance of Aerobic Denitrification SynCom
| Parameter | Undisturbed Performance | DBP Disturbance | LOFX Disturbance | Measurement Method |
|---|---|---|---|---|
| NO₃⁻-N Removal Efficiency | 94.0% ± 3.3% | 93.1% ± 2.7% | 92.8% ± 3.1% | Spectrophotometric analysis |
| AHL Signaling Molecules | C6-HSL, 3OC6-HSL dominant | C4-HSL dominant | 3OC12-HSL dominant | LC-MS/MS quantification |
| Electron Transfer Activity | Baseline (100%) | 142% ± 15% increase | 127% ± 12% increase | Cyclic voltammetry |
| TCA Cycle Metabolites | Normal flux | 2.1-fold increase | 1.8-fold increase | Metabolomic profiling |
| EPS Production | 125.3 mg/L ± 12.4 | 283.7 mg/L ± 24.6 | 231.5 mg/L ± 19.8 | Phenol-sulfuric acid method |
Protocol: Stability Assessment Under Environmental Stress
Materials:
- Established synthetic community
- Disturbance agents (e.g., antibiotics, pollutants)
- LC-MS/MS for AHL quantification
- Electrochemical workstation for electron transfer measurement
- RNA extraction kit and sequencing platform for metatranscriptomics
Procedure:
- Community Acclimation: Grow SynCom in appropriate medium for 48 hours to establish stable interactions.
- Disturbance Application: Introduce sublethal concentrations of disturbance agents (e.g., 200 μg/L SMX, environmental concentrations of DBP/LOFX).
- Functional Monitoring: Track key community functions (e.g., denitrification efficiency, biomass production) throughout disturbance period.
- Molecular Analysis: Quantify signaling molecules (AHLs) via LC-MS/MS at 12-hour intervals.
- Electron Transfer Measurement: Assess electron transfer activity using cyclic voltammetry.
- Metatranscriptomic Profiling: Sequence community RNA to identify metabolic pathway regulation.
- Metabolic Network Reconstruction: Integrate omics data to map functional adaptations.
Figure 2: Stability Maintenance Pathways Under Environmental Disturbance. SynComs maintain function through coordinated molecular, metabolic, and ecological responses to stress [21].
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for SynCom Stability Studies
| Reagent/Category | Function/Application | Example Specifications | Experimental Use Cases |
|---|---|---|---|
| AHL Standards | Quantification of quorum sensing molecules | C4-HSL, C6-HSL, 3OC12-HSL (Sigma-Aldrich) | LC-MS/MS calibration for interspecific communication monitoring [21] |
| Antibiotic Resistance Plasmids | Engineering cooperator-cheater dynamics | pFPV-LuxR, pFPV-Sul1 in P. aeruginosa [57] | Constructing detoxifying cooperators and sensitive cheaters for social dynamics studies |
| Selective Media Components | Tracking individual strain dynamics | SMX (10 μg/mL), gentamycin (30 μg/mL) supplementation [57] | Differential colony counting of cooperator vs. cheater strains in coculture |
| Metabolic Probes | Monitoring pathway activity | 13C-labeled substrates for flux analysis | Quantifying metabolic rewiring and cross-feeding interactions |
| Microplate Assay Kits | High-throughput function screening | INT, CTC for electron transfer activity | Community functional screening in factorial designs [59] |
| DNA/RNA Preservation Buffers | Multi-omics sample preparation | RNAlater, DNA/RNA Shield | Preservation of community samples for metatranscriptomic and metagenomic analysis |
Stabilizing synthetic microbial communities against cheater invasion and competitive collapse requires integrated strategies spanning spatial engineering, metabolic network design, and disturbance-responsive regulation. The protocols and mechanisms outlined here provide a roadmap for constructing robust SynComs that maintain functional stability in bioproduction, bioremediation, and therapeutic applications. By leveraging full factorial construction to map community-function landscapes and implementing stability mechanisms informed by natural community principles, researchers can design synergistic microbial consortia resistant to social cheating and environmental perturbations.
Balancing Diversity and Functionality to Avoid Cooperation Breakdown
Synthetic Microbial Communities (SynComs) represent a paradigm shift in biotechnology, enabling complex metabolic functions that are challenging to engineer into single strains [2]. However, a critical challenge persists: cooperation breakdown due to the emergence of cheater strains that exploit community resources without contributing to collective functionality [2]. This protocol addresses the fundamental trade-off between diversity and functionality, providing a structured framework for designing stable, high-performance consortia through comparative metabolic modeling. The instability primarily stems from metabolic cheating, where non-productive members gain fitness advantages by avoiding metabolic costs of cooperative functions, ultimately leading to community collapse [2]. By integrating ecological principles with computational modeling, we establish methodologies to preemptively identify and mitigate these failure modes, enabling robust SynCom design for biomedical, bioproduction, and environmental applications.
Computational Design Protocols
Genome-Scale Metabolic Model (GEM) Reconstruction
Objective: Build organism-specific metabolic networks to serve as foundation for community modeling.
Protocol Steps:
- Draft Reconstruction: Generate an initial model from genomic annotations using automated pipelines (ModelSEED, RAVEN, or COBRA Toolbox) [26] [60]. Map gene-protein-reaction (GPR) associations to establish metabolic capabilities.
- Network Curation: Manually refine the draft model using organism-specific biochemical literature and experimental data [26]. Critical curation steps include:
- Defining reaction directionality based on thermodynamic constraints
- Incorporating substrate and cofactor specificity of enzymes
- Adding transport reactions for nutrient uptake and metabolite secretion
- Validating growth capabilities under different nutrient conditions
- Gap Filling: Identify and fill metabolic gaps that prevent synthesis of essential biomass precursors [26]. Prioritize reactions with genomic evidence over those added solely to enable metabolic functionality.
- Biomass Equation Formulation: Define a comprehensive biomass equation representing cellular composition, including amino acids, nucleotides, lipids, and cofactors, weighted by experimentally measured proportions [26].
- Model Validation: Test the model's predictive accuracy against experimental data, including gene essentiality, substrate utilization patterns, and byproduct secretion profiles [60]. Iteratively refine until >90% accuracy is achieved for essential gene predictions.
Quality Control: The reconstruction process should adhere to established standards for high-quality GEMs, with complete documentation of all data sources and curation decisions [26].
Community-Level Model Integration
Objective: Integrate individual GEMs into a community metabolic model that simulates interspecies interactions.
Protocol Steps:
- Model Compartmentalization: Maintain individual organism GEMs as separate compartments within the community model [23]. Create a shared extracellular environment compartment for metabolite exchange.
- Metabolite Exchange Network: Define the set of metabolites that can be transferred between community members and their external environment [23]. This includes:
- Cross-fed metabolites (e.g., amino acids, vitamins, metabolic byproducts)
- Public goods (e.g., enzymes, siderophores)
- Waste products that may serve as nutrients for other members
- Interaction Framework Selection: Choose an appropriate modeling framework based on the community complexity and available data:
- Constraint Definition: Apply constraints to represent ecological realities:
- Nutrient availability limits based on environmental conditions
- Thermodynamic constraints on reaction fluxes
- Capacity constraints on transport reactions
Table 1: Metabolic Modeling Approaches for Microbial Communities
| Model Type | Best Application Context | Key Advantages | Limitations |
|---|---|---|---|
| Compartmentalized Static (FBA) | Defined synthetic consortia with balanced diversity [23] | High prediction accuracy for well-characterized systems; Computationally efficient | Requires detailed species-specific data |
| Compartmentalized Dynamic | Communities with strong temporal dynamics [23] | Captures population shifts over time; Models succession patterns | Requires extensive parameter estimation |
| Lumped Network | Complex natural communities [23] | Works with metagenomic data only; Estimates community metabolic potential | Overestimates capabilities; Loses species resolution |
| Multi-Objective (OptCom) | Systems with clear individual/community trade-offs [23] | Captures altruistic/selfish interactions; Models evolutionary conflicts | Computationally intensive; Complex implementation |
The following diagram illustrates the workflow for constructing and simulating community metabolic models:
Workflow for Community Metabolic Modeling
Stability and Cheating Risk Assessment
Objective: Identify potential instability drivers and cheater formation risks in silico before experimental implementation.
Protocol Steps:
- Flux Variability Analysis: Determine the range of possible metabolic fluxes for each reaction in the network under community conditions [23]. Large variabilities in public goods production indicate potential cheating vulnerabilities.
- Single-Gene Deletion Studies: Systematically knock out each gene in the model to identify:
- Metabolic dependencies between community members
- Essential functions that could become targets for exploitation
- Reactions whose disruption creates cheater phenotypes
- Sensitivity Analysis: Perturb nutrient inputs and environmental conditions to identify parameter regions where community stability breaks down [2]. Pay particular attention to:
- Carbon-to-nitrogen ratios that trigger competitive exclusion
- Oxygen gradients that create niche specialization
- pH fluctuations that differentially affect member growth
- Cheater Prediction: Identify strains that can survive on community-produced metabolites without contributing to collective functions [2]. Flag strains that:
- Lack biosynthetic pathways for essential metabolites but possess uptake systems
- Show minimal investment in public goods production
- Maximize growth yield when metabolic costs are offloaded to other members
Experimental Validation Protocols
Community Assembly and Monitoring
Objective: Construct predicted stable communities and validate their functionality and stability over time.
Protocol Steps:
- Strain Preparation: Cultivate individual community members in isolation, then combine in proportions predicted by metabolic modeling [2]. Include control communities with known instability patterns for comparison.
- Longitudinal Monitoring: Track community composition and function over multiple generations (≥50) using:
- Flow cytometry: With strain-specific tagging for population quantification
- qPCR: Targeting species-specific genetic markers
- Metabolite profiling: HPLC/MS to measure metabolic cross-feeding
- Function-specific assays: Measuring target output (e.g., product formation, substrate degradation)
- Perturbation Response Testing: Subject communities to defined disturbances to assess resilience:
- Nutrient pulsing or starvation cycles
- Invasion by cheat strains or external species
- Antibiotic exposure at sub-inhibitory concentrations
- pH or temperature shifts
- Spatial Structure Evaluation: For communities intended for biofilm or surface-associated applications, test stability in both well-mixed and spatially structured environments [2]. Spatial structure often suppresses cheating by creating physical barriers to exploit public goods [2].
Table 2: Research Reagent Solutions for Experimental Validation
| Reagent/Category | Specific Examples | Function in Protocol |
|---|---|---|
| Fluorescent Tags | GFP, RFP, mCherry variants | Strain-specific labeling for population tracking |
| Selection Markers | Antibiotic resistance genes | Maintain engineered functions; selective pressure |
| Quorum Sensing Systems | LuxI/LuxR, LasI/LasR | Engineered communication for coordination |
| Bacteriocins | MccV, nisin [61] | Targeted growth inhibition for stability |
| Culture Systems | Chemostats, microfluidics devices | Maintain steady-state conditions; spatial structure |
| Reporter Systems | Transcriptional fusions, biosensors | Monitor gene expression and metabolite production |
Cheater Detection and Mitigation
Objective: Identify and suppress cheater emergence in experimental communities.
Protocol Steps:
- Cheater Isolation: Periodically isolate community members and test their individual capabilities to identify functional losses [2]. Compare isolated strains to ancestors through:
- Metabolic profiling of substrate utilization
- Genome resequencing to identify mutations
- Measurement of public goods production in monoculture
- Spatial Containment: Implement physical barriers to cheater spread through:
- Encapsulation in hydrogel beads or microfluidic droplets
- Membrane-based compartmentalization
- Biofilm cultivation on structured surfaces
- Evolutionary Steering: Apply selective pressure that favors cooperative phenotypes:
- Link essential nutrient access to cooperative function output
- Implement negative selection against non-producers using toxin-antitoxin systems [61]
- Periodic environment switching that favors different functional groups
- Mathematical Verification: Fit experimental data to ecological models to quantify interaction strengths and identify destabilizing dynamics [2]. Use parameters from metabolic models to inform these ecological models.
Implementation Guidelines
Diversity-Functionality Optimization
Objective: Balance taxonomic and functional diversity to maximize performance while minimizing instability.
Protocol Steps:
- Keystone Identification: Use metabolic models to identify potential keystone species that disproportionately impact community function through:
- Central positioning in metabolic interaction networks
- Production of public goods with broad community benefits
- Functional roles with low redundancy across the community
- Modular Design: Structure communities with functional modules containing metabolically interdependent species [2]. Implement "division of labor" where different modules handle distinct metabolic tasks to reduce direct competition [2].
- Interaction Engineering: Strategically balance positive and negative interactions:
- Functional Redundancy: Maintain multiple species capable of performing critical functions while minimizing direct metabolic overlap that creates intense competition [2].
The following diagram illustrates the key principles for maintaining community stability:
Principles for Community Stability
Model-Experiment Integration
Objective: Establish iterative cycles between computational prediction and experimental validation.
Protocol Steps:
- Design-Build-Test-Learn (DBTL) Implementation: Follow structured iteration:
- Multi-Omics Data Integration: Incorporate experimental data to constrain and validate models:
- Metatranscriptomics: Identify actively expressed pathways
- Metaproteomics: Quantify enzyme abundance and metabolic investments
- Metabolomics: Measure metabolite exchange fluxes and identify cross-fed compounds
- Parameter Refinement: Use experimental data to improve model accuracy by:
- Adjusting maintenance energy requirements based on measured growth rates
- Constraining uptake rates using measured nutrient depletion profiles
- Refining biomass equations based on elemental composition analysis
- Stability Metric Development: Establish quantitative measures for community stability:
- Resistance: Magnitude of functional change after perturbation
- Resilience: Speed of functional recovery after perturbation
- Robustness: Maintenance of function across environmental variations
This protocol establishes a comprehensive framework for balancing diversity and functionality in synthetic microbial communities while preventing cooperation breakdown. By integrating comparative metabolic modeling with experimental validation, we enable predictive design of stable, high-performance consortia. The key innovation lies in preemptively identifying and mitigating cheating risks through computational analysis before experimental implementation, significantly reducing development time and resource investment. As the field advances, integration of machine learning with multi-scale models will further enhance our ability to design complex microbial ecosystems with precisely controlled functions and stability.
Spatial Engineering and Modular Design to Enhance Coexistence and Function
Application Note: Computational Design of Synthetic Microbial Consortia
Synthetic microbial consortia represent a paradigm shift in biotechnology, enabling complex functions through division of labor among microbial subpopulations. Unlike monocultures, consortia can perform more complex tasks, utilize simpler substrates, and exhibit increased robustness to environmental perturbations [62]. Spatial engineering and modular design are critical for stabilizing these communities against competitive exclusion and optimizing their functional output. This application note details integrated computational and experimental workflows for designing, modeling, and implementing robust synthetic microbial ecosystems, with a specific focus on leveraging comparative metabolic modeling to predict and enhance community coexistence and productivity.
Computational Workflow for Community Design
The design of stable synthetic communities follows a structured iterative cycle, integrating computational modeling with experimental validation. The core workflow involves two primary stages: in silico system design and experimental implementation and analysis.
The following diagram illustrates the integrated computational-experimental workflow for designing synthetic microbial communities, from initial conceptualization to final analysis.
Key Computational Protocols
Protocol 1.1: Reconstructing Consensus Genome-Scale Metabolic Models (GEMs) for Community Members
Purpose: To generate high-quality, predictive metabolic models for each member of the proposed microbial consortium. Consensus approaches that integrate multiple reconstruction tools have been shown to produce more comprehensive and functional models [27].
- Step 1: Data Acquisition. Obtain high-quality genomic data for the target microbial strains. For complex communities, this may involve extracting Metagenome-Assembled Genomes (MAGs) [27].
- Step 2: Draft Reconstruction. Use automated reconstruction tools (e.g., CarveMe, gapseq, KBase) to generate draft metabolic models from the genomic data. CarveMe uses a top-down approach with a universal template, while gapseq and KBase employ bottom-up strategies [27].
- Step 3: Build Consensus Model. Employ a consensus pipeline to merge the draft models from the different tools. This integration results in a model that includes a larger number of reactions and metabolites while reducing the number of non-functional dead-end metabolites [27].
- Step 4: Network Refinement and Gap-Filling. Use a constraint-based gap-filling algorithm like COMMIT to ensure the model can produce essential biomass precursors under a defined minimal medium. This step adds necessary reactions to restore metabolic functionality [27].
- Step 5: Quality Control. Validate the model's functionality by testing its ability to simulate known metabolic capabilities, such as growth on specific carbon sources or the production of key metabolites.
Protocol 1.2: Automated Model Selection for Stable Community Design (AutoCD)
Purpose: To computationally generate and rank all possible two- or three-strain community designs based on their probability of achieving a stable, steady-state coexistence [61].
- Step 1: Define the Part Library. Specify the available biological parts: number of strains (N), bacteriocins (B), and quorum sensing (QS) systems (A). Define how QS systems can regulate bacteriocin expression (induction/repression) [61].
- Step 2: Generate Candidate Model Space. A model space generator creates all unique combinations of strains expressing different genetic circuits. Each candidate model includes nutrient-based competition and engineered interactions (e.g., QS-regulated bacteriocin production) [61].
- Step 3: Define Objective Behavior. Mathematically define the target behavior—a stable steady state. This is typically described using distance functions that quantify how far a simulation is from stability (e.g., based on population gradients, standard deviations, and minimum population densities) [61].
- Step 4: Perform Model Selection. Use Approximate Bayesian Computation with Sequential Monte Carlo (ABC SMC) to sample models and parameters from prior distributions. The algorithm iteratively propagates particles, gradually refining distance thresholds until they meet the objective criteria, providing a posterior probability for each candidate model [61].
- Step 5: Identify Optimal Designs. The model with the highest posterior probability (e.g., a cross-protection mutualism system where strains reciprocally repress self-limiting bacteriocins) represents the most robust design for experimental implementation [61].
Protocol 1.3: Comparing Metabolic States with ComMet
Purpose: To identify condition-specific metabolic features and potential interaction motifs by comparing the flux spaces of community GEMs under different constraints [63].
- Step 1: Define Metabolic States. Set up the GEMs to represent different metabolic states of the community (e.g., unlimited vs. limited nutrient uptake, different strain ratios) by applying specific constraints to exchange reactions.
- Step 2: Analyze Flux Spaces. Instead of using Flux Balance Analysis (FBA), which requires a predefined objective function, use the ComMet method. It combines an analytical approximation of flux probability distributions with Principal Component Analysis (PCA) to characterize the feasible flux space without the need for computationally intensive sampling [63].
- Step 3: Extract Discriminative Modules. Perform PCA on the flux space to decompose it into biochemically interpretable reaction sets, or "modules." Identify the modules whose flux variability accounts for the major differences between the metabolic states [63].
- Step 4: Visualize and Interpret. Map the discriminative modules onto metabolic network maps to visualize the underlying functional differences, such as alterations in TCA cycle activity or fatty acid metabolism, which can inform on potential synergistic or competitive interactions within the community [63].
Application Note: Experimental Implementation of Spatially Structured Communities
Spatial Engineering Platforms
Overcoming the challenge of competitive exclusion in a shared environment requires engineering the habitat itself. Spatially Linked Microbial Consortia (SLMC) provide a physical framework to control interactions and optimize local conditions for each member [62].
The following diagram outlines the architecture of a Spatially Linked Microbial Consortia (SLMC) platform, showing how separate modules are connected to control metabolic exchanges.
Key Experimental Protocols
Protocol 2.1: Establishing a Spatially Linked Microbial Consortia (SLMC) in a Bioreactor
Purpose: To physically separate interacting microbial strains into distinct modules with independently optimized environmental conditions, while enabling controlled metabolic exchanges between them [62].
- Step 1: Module Configuration. Set up multiple bioreactor vessels or hollow-fiber modules. Each module will host a single microbial strain or a defined sub-community.
- Step 2: Environmental Optimization. Independently control and optimize critical environmental parameters (e.g., pH, temperature, dissolved oxygen, medium composition) within each module to suit the specific needs of the resident strain(s) [62].
- Step 3: System Interlinking. Connect the modules via a controlled perfusion system that regulates the flux of media and metabolites between them. This can be achieved using pumps for convective flow or membranes for diffusive exchange [62].
- Step 4: Inoculation and Operation. Inoculate individual strains into their respective modules. Initiate the controlled exchange of media between modules to establish metabolic communication.
- Step 5: Monitoring. Regularly sample from each module to monitor population density (e.g., via flow cytometry or OD measurements) and metabolite concentrations (e.g., via HPLC or MS) to track community dynamics and function.
Protocol 2.2: Dynamic Visualization of Metabolic States using GEM-Vis
Purpose: To create animated visualizations of time-course metabolomic data within the context of a metabolic network, providing an intuitive tool for analyzing community dynamics and metabolic interactions [64].
- Step 1: Data and Model Preparation. Obtain a manually drawn or algorithmically generated metabolic network map (e.g., in SBML format) and a quantitative time-course metabolomic data set [64].
- Step 2: Data Integration. Use the SBMLsimulator software to load the model and data. Map the measured metabolites to their corresponding nodes in the network.
- Step 3: Animation Setup. In SBMLsimulator, select the "fill level" of each metabolite node as the visual representation of its concentration over time. This method allows for the most intuitive estimation of quantitative changes by human observers [64].
- Step 4: Generate Animation. Run the software to create a smooth animation interpolating between the measured time points. The resulting video shows the dynamic changes in the metabolic network, highlighting pathways that are activated or deactivated over the course of the experiment [64].
- Step 5: Analysis. Use the animation to generate hypotheses about metabolic interactions, such as the coordinated accumulation of metabolites in linked pathways, which may not be apparent from static data analysis [64].
The Scientist's Toolkit: Research Reagent Solutions
Table 1: Essential Computational Tools and Databases for Metabolic Modeling of Microbial Communities
| Tool Name | Type | Primary Function | Relevance to Community Modeling |
|---|---|---|---|
| CarveMe [27] | Software Tool | Automated, top-down GEM reconstruction | Fast generation of draft models from genome annotations. |
| gapseq [27] | Software Tool | Automated, bottom-up GEM reconstruction | Comprehensive biochemical network inference from genomic data. |
| KBase [27] | Software Platform | Integrated GEM reconstruction and analysis | User-friendly platform for model building and simulation. |
| COBRA Toolbox [26] [65] | Software Suite | Constraint-Based Modeling and Analysis | Essential suite for simulation, gap-filling, and analyzing GEMs (e.g., FBA, sampling). |
| COMMIT [27] | Algorithm | Community Model Gap-Filling | Gap-filling metabolic models in a community context to ensure metabolic functionality. |
| ModelSEED [27] | Biochemical Database | Reaction Database for GEMs | Standardized biochemical database used by tools like gapseq and KBase. |
| AutoCD [61] | Computational Workflow | Automated Community Design | Generates and ranks candidate community interaction networks for stability. |
| ComMet [63] | Computational Method | Comparison of Metabolic States | Identifies differential metabolic features between community states without a predefined objective function. |
| MetaboTools [65] | Toolbox | Analysis of GEMs with Omics Data | Facilitates integration of extracellular metabolomic data into GEMs for contextualized analysis. |
| GEM-Vis [64] | Visualization Method | Dynamic Visualization of Metabolomic Data | Creates animations of time-course metabolomic data on network maps for intuitive analysis. |
Table 2: Key Experimental Platforms and Biological Parts for Spatial Engineering
| Reagent / Platform | Type | Primary Function | Relevance to Spatial Engineering |
|---|---|---|---|
| Spatially Linked Bioreactor (SLMC) [62] | Hardware Platform | Modular Cultivation System | Enables physical separation of strains with controlled metabolic exchange under optimized per-strain conditions. |
| Hollow-Fiber Bioreactor [62] | Hardware Platform | Membrane-based Co-culture | Allows diffusive exchange of small molecules between spatially segregated populations. |
| Quorum Sensing (QS) Systems [61] | Biological Part | Cell-Cell Communication Module | Engineered for regulating bacteriocin or metabolic gene expression in response to population density. |
| Bacteriocins [61] | Biological Part | Amensal Interaction Module | Toxins (e.g., MccV, nisin) used to selectively manipulate growth rates of sensitive subpopulations and stabilize communities. |
| SBMLsimulator [64] | Software Tool | Dynamic Model Simulation & Visualization | Used in conjunction with GEM-Vis to create animations of dynamic metabolic network data. |
From In Silico to In Vivo: Validating Predictive Models and Benchmarking Performance
The reconstruction of genome-scale metabolic models (GEMs) is a fundamental process in systems biology, enabling the in silico study of metabolic capabilities in microbial communities. Multiple automated reconstruction tools are available, yet they produce models with significant structural and functional variations. This application note provides a standardized protocol for benchmarking these tools, with a focus on Jaccard similarity analysis of reactions, metabolites, and genes. We demonstrate that consensus approaches mitigate tool-specific biases and enhance model accuracy for synthetic microbial community research, supported by quantitative comparisons and detailed experimental workflows.
Genome-scale metabolic models (GEMs) serve as powerful computational frameworks for predicting metabolic behaviors and interactions in synthetic microbial communities (SynComs). The accuracy of these predictions, however, depends heavily on the quality of the reconstructed models [13]. Several automated reconstruction tools—including CarveMe, gapseq, and KBase—have been developed, each utilizing distinct biochemical databases and algorithms [13]. This diversity leads to substantial variations in model content, including the sets of reactions, metabolites, and genes incorporated.
The Jaccard similarity index has emerged as a critical metric for quantifying these differences, providing a standardized measure of overlap between sets of model components [13]. For the comparative metabolic modeling of synthetic microbial communities, such benchmarking is essential to ensure reliable predictions of metabolic interactions and community functions. This protocol details a comprehensive framework for evaluating reconstruction tools, emphasizing Jaccard-based comparisons and the development of consensus models that integrate strengths across multiple tools.
Comparative Analysis of Model Structures
Reconstruction tools applied to the same genomic input can generate GEMs with markedly different structural properties. Understanding these differences is a prerequisite for effective tool selection and consensus modeling.
Quantitative Structural Variations
A comparative analysis of GEMs reconstructed from the same metagenome-assembled genomes (MAGs) using CarveMe, gapseq, and KBase reveals significant disparities in model composition. The following table summarizes the structural characteristics observed from two marine bacterial communities [13].
Table 1: Structural Characteristics of GEMs from Different Reconstruction Tools
| Reconstruction Tool | Number of Reactions | Number of Metabolites | Number of Genes | Number of Dead-End Metabolites |
|---|---|---|---|---|
| gapseq | Highest | Highest | Lowest | Highest |
| CarveMe | Intermediate | Intermediate | Highest | Intermediate |
| KBase | Intermediate | Intermediate | Intermediate | Lowest |
| Consensus | High | High | High | Reduced |
Jaccard Similarity of Model Components
The Jaccard similarity coefficient, which measures the overlap between two sets, was calculated for reactions, metabolites, and genes from models derived from the same MAGs. The analysis demonstrated low to moderate similarity across all components, underscoring the tool-specific biases [13].
Table 2: Average Jaccard Similarity Between Reconstruction Tools
| Compared Tools | Reactions | Metabolites | Genes |
|---|---|---|---|
| gapseq vs KBase | 0.23-0.24 | 0.37 | 0.42-0.45 |
| gapseq vs CarveMe | Low | Low | Low |
| CarveMe vs KBase | Low | Low | Moderate |
| CarveMe vs Consensus | - | - | 0.75-0.77 |
The notably higher similarity between CarveMe and consensus models in gene content suggests that consensus approaches effectively integrate genomic evidence from multiple sources, thereby reducing the bias inherent in any single tool [13].
Experimental Protocols
This section provides a detailed, step-by-step protocol for reconstructing and benchmarking GEMs, from data preparation to Jaccard similarity analysis.
Genome-Scale Metabolic Model Reconstruction
Objective: To reconstruct draft GEMs from genomic data using three automated tools (CarveMe, gapseq, and KBase).
Materials and Reagents:
- Genomic Input: High-quality MAGs or isolate genomes in FASTA format.
- Software Tools: CarveMe (v1.5.1), gapseq (v1.3.1), KBase (Narrative interface).
- Computing Environment: Unix-based system (for CarveMe and gapseq), web browser (for KBase), and R (v4.0.0+) for downstream analysis.
Procedure:
- Input Preparation: Ensure your genomic data (MAGs) are in FASTA format and note their file paths.
- Tool Execution:
- CarveMe: Use the following command to reconstruct a model for each MAG.
- gapseq: Execute the
gapseqpipeline to generate a metabolic model. - KBase: Upload genomes to the KBase platform and use the "Build Metabolic Model" app in the Narrative interface.
- Model Standardization: Convert all output models to a common SBML format using available conversion scripts (e.g.,
cobrapyin Python) to ensure compatibility for comparison. - Data Extraction: Parse the SBML files to extract comprehensive lists of reactions, metabolites, and genes for each generated model. Store these as sets for subsequent similarity analysis.
Jaccard Similarity Calculation
Objective: To quantitatively assess the pairwise similarity of the models generated by different tools.
Theory: The Jaccard similarity coefficient between two sets A and B is calculated as: Jaccard(A, B) = |A ∩ B| / |A ∪ B|
The value ranges from 0 (no overlap) to 1 (identical sets).
Procedure:
- For each pair of tools (e.g., Tool A and Tool B) and for each MAG, extract the three sets:
- Set of Reactions (R)
- Set of Metabolites (M)
- Set of Genes (G)
- Calculate the three Jaccard indices for each model component.
- JaccardR = |RA ∩ RB| / |RA ∪ RB|
- JaccardM = |MA ∩ MB| / |MA ∪ MB|
- Jaccard_G = |GA ∩ GB| / |GA ∪ GB|
- Repeat this calculation for all MAGs in the dataset and compute the average Jaccard similarity for each component pair across the community.
Consensus Model Generation
Objective: To integrate multiple draft reconstructions into a single, more comprehensive consensus model.
Procedure:
- Model Aggregation: For a given MAG, combine the reactions, metabolites, and genes from the GEMs generated by CarveMe, gapseq, and KBase.
- Reaction Inclusion: A reaction is included in the draft consensus model if it appears in at least two of the three individual reconstructions [13].
- Gap-Filling: Use the COMMIT tool to perform network gap-filling on the draft consensus model. This step adds minimal reactions to ensure network functionality and connectivity within a defined medium [13].
Functional Validation
Objective: To validate the predictive power of the original and consensus models.
Procedure:
- Gene Essentiality Prediction: Use Flux Balance Analysis (FBA) with each model to predict essential genes. Set biomass production as the objective function and simulate the knockout of each gene.
- Benchmarking against Experimental Data: Compare the in silico essentiality predictions with experimental data from shRNA knockout screens [66]. Calculate the enrichment of predicted essential genes in the experimentally essential gene set.
- Metabolite Exchange Prediction: Simulate the community metabolic model and predict the set of exchanged metabolites. Compare these predictions with experimentally measured exchange profiles, if available.
The Scientist's Toolkit
Table 3: Essential Research Reagents and Computational Tools
| Item Name | Function/Application | Specifications |
|---|---|---|
| CarveMe | Top-down GEM reconstruction from genomes. Uses a universal model template. | Speed: Fast; Template: AUROME [13]. |
| gapseq | Bottom-up GEM reconstruction using extensive biochemical databases. | Database: Comprehensive; Approach: Evidence-based [4] [13]. |
| KBase | Web-based, user-friendly platform for systems biology analysis. | Approach: Bottom-up; Database: ModelSEED [13]. |
| COMMIT | Community Model Integration Tool for gap-filling metabolic networks. | Application: Gap-filling consensus models [13]. |
| BacArena | Tool for dynamic simulation of microbial communities using GEMs. | Application: Simulating SynCom dynamics [4]. |
| GapSeq | Metabolic pathway prediction and GEM reconstruction tool. | Application: Generates models compatible with BacArena [4]. |
| HMSC | Host-Microbe Systems Biology framework for integrative modeling. | Application: Studying host-microbe interactions [12]. |
Workflow Visualization
The following diagram illustrates the complete experimental and computational workflow for benchmarking reconstruction tools and building consensus models for synthetic communities.
Figure 1: Workflow for benchmarking GEM reconstruction tools. The process begins with Metagenome-Assembled Genomes (MAGs), progresses through parallel reconstruction with different tools, and culminates in quantitative comparison and consensus model generation.
Benchmarking metabolic reconstruction tools through Jaccard similarity analysis is a critical step in developing reliable models for synthetic microbial communities. The quantitative data presented herein clearly demonstrate that tool selection significantly influences model structure and content. The consensus reconstruction approach mitigates individual tool biases and generates more comprehensive models, as evidenced by higher gene content similarity and reduced dead-end metabolites. This protocol provides a standardized framework for researchers to critically evaluate and integrate GEMs, thereby enhancing the predictive accuracy of metabolic interactions in engineered microbial ecosystems.
The rational engineering of synthetic microbial consortia for therapeutic and biotechnological applications requires precise understanding and prediction of community metabolic fluxes. While genome-scale metabolic models (GEMS) provide powerful computational frameworks for predicting metabolic fluxes in silico, their predictive accuracy hinges on validation against experimental data [36]. Quantitative proteomic data provides a crucial link between model predictions and cellular physiology by quantifying the abundance of metabolic enzymes that catalyze flux-carrying reactions [67]. This Application Note details integrated methodologies for correlating proteomic profiling with metabolic modeling outputs to validate and refine flux predictions in synthetic microbial communities, thereby enhancing the predictive power of in silico models for therapeutic development.
Foundational Concepts
Metabolic Modeling of Microbial Communities
Genome-scale metabolic modeling employs mathematical representations of biochemical reaction networks to predict metabolic capabilities and behaviors. For microbial communities, three primary modeling approaches are commonly utilized, each with distinct advantages and limitations [68] [27]:
- Compartmentalized Modeling: Individual GEMs are merged into a single stoichiometric matrix with a shared extracellular compartment, enabling metabolite exchange while maintaining species-specific reaction spaces [68].
- Lumped Model (Mixed-Bag): All metabolic reactions from community members are pooled into a single model with one cytosolic compartment, treating the community as a supra-organism [68] [27].
- Costless Secretion: Models are simulated separately while dynamically updating the shared environment based on metabolites secreted without growth cost [68].
Constraint-based analysis techniques, including Flux Balance Analysis (FBA) and Flux Sampling, are applied to these model structures to predict metabolic flux distributions. FBA identifies optimal flux distributions that maximize a cellular objective (typically biomass production), while flux sampling uses Markov chain Monte Carlo methods to randomly generate thermodynamically-feasible flux distributions without presupposing a cellular objective, thereby exploring phenotypic heterogeneity and reducing user-introduced bias [68].
Quantitative Proteomics for Systems Pharmacology
Mass spectrometry-based proteomics enables large-scale quantification of proteins within biological systems. Two primary approaches are employed in translational pharmacology [67]:
- Untargeted (Global) Proteomics: Discovers and identifies proteins across a broad spectrum without prior selection, ideal for hypothesis generation and comprehensive system characterization.
- Targeted Proteomics: Quantifies predefined proteins with high reproducibility, precision, and sensitivity, using stable isotope-labeled standards for absolute quantification. This approach is essential for validating specific protein targets.
Quantitative proteomic data directly informs in vitro-in vivo extrapolation (IVIVE) within physiologically-based pharmacokinetic (PBPK) models by providing absolute abundance values for key proteins involved in drug absorption, distribution, metabolism, and excretion (ADME) [67]. When applied to microbial communities, these data serve as critical constraints for metabolic models, tethering in silico predictions to experimentally measurable cellular components.
Integrated Protocol for Correlation Analysis
This protocol outlines a systematic workflow for acquiring proteomic data and correlating it with flux predictions from metabolic models of synthetic microbial communities.
Proteomic Profiling of Microbial Communities
Method: Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS) for Protein Quantification
Procedure:
Sample Preparation:
- Culture synthetic microbial communities under defined conditions.
- Harvest cells at mid-exponential growth phase by rapid centrifugation (4,000 x g, 10 min, 4°C).
- Lyse cells using RIPA buffer supplemented with protease inhibitors.
- Reduce disulfide bonds with 5 mM dithiothreitol (37°C, 30 min) and alkylate with 10 mM iodoacetamide (room temperature, 30 min in darkness).
- Digest proteins with sequencing-grade trypsin (1:50 enzyme-to-protein ratio) at 37°C for 16 hours [69].
LC-MS/MS Analysis:
- Desalt digested peptides using C18 solid-phase extraction columns.
- Separate peptides via reverse-phase C18 nano-liquid chromatography using a 60-minute gradient from 2% to 35% acetonitrile in 0.1% formic acid.
- Analyze eluted peptides using a high-resolution tandem mass spectrometer operating in data-dependent acquisition mode for untargeted discovery, or parallel reaction monitoring (PRM) mode for targeted quantification [70].
Data Processing:
- Identify proteins by searching MS/MS spectra against a curated database containing protein sequences from all consortium members.
- For absolute quantification using targeted proteomics, use stable isotope-labeled standard peptides for calibration [67]. Generate calibration curves for each target protein and calculate absolute abundances in fmol/μg total protein.
Table 1: Key Research Reagents for Proteomic Profiling
| Reagent/Category | Specific Examples | Function in Protocol |
|---|---|---|
| Digestion Enzyme | Sequencing-grade trypsin | Proteolytic cleavage of proteins into peptides for MS analysis [69] |
| Reducing Agent | Dithiothreitol (DTT) | Reduction of protein disulfide bonds |
| Alkylating Agent | Iodoacetamide | Cysteine alkylation to prevent reformation of disulfide bonds |
| Chromatography Column | Reverse-phase C18 nano-column | Peptide separation by hydrophobicity |
| Mass Spectrometer | High-resolution tandem MS | Peptide identification and quantification [70] |
| Isotope Standards | Stable isotope-labeled standard peptides | Absolute quantification of target proteins [67] |
Metabolic Model Reconstruction and Flux Prediction
Method: Constraint-Based Reconstruction and Analysis (COBRA)
Procedure:
Community Model Reconstruction:
- Obtain genome sequences for all microbial consortium members.
- Reconstruct draft GEMs using automated tools such as CarveMe, gapseq, or KBase. Consider constructing consensus models from multiple tools to reduce reconstruction bias and increase metabolic coverage [27].
- For consortium modeling, implement a compartmentalized approach, combining individual GEMs into a single model with a shared extracellular space [68].
Flux Prediction:
- Constrain the model with experimentally measured substrate uptake rates.
- Perform flux sampling using algorithms such as Constrained Riemannian Hamiltonian Monte Carlo to generate a statistically representative set of feasible flux distributions. Use 1000 samples per simulation to adequately capture flux space [68].
- Alternatively, perform parsimonious FBA to obtain a single, optimal flux distribution for comparison.
Data Integration and Correlation Analysis
Method: Proteomic-Constraint Integration and Statistical Correlation
Procedure:
Proteomic Data Integration:
- Map quantified metabolic enzymes to their corresponding gene-protein-reaction (GPR) associations in the GEM.
- Convert absolute protein abundances (fmol/μg) into relative abundance measures normalized to the sum of all quantified enzymes.
- Implement a metabolic constraint that weights reaction flux capacities by the relative abundance of the catalyzing enzyme(s).
Correlation Analysis:
- Extract median flux values for each reaction from the sampled flux distributions.
- Calculate Spearman correlation coefficients between median reaction fluxes and the relative abundances of their corresponding enzyme complexes.
- Statistically significant correlations (p-value < 0.05, adjusted for multiple testing) indicate reactions where proteomic data validates model predictions.
- For reactions showing poor correlation (p-value > 0.05), investigate potential gaps in model annotation, post-translational regulation, or measurement errors in proteomic data.
Table 2: Representative Correlation Data Between Enzyme Abundance and Predicted Flux
| Reaction ID | Enzyme Complex | Protein Abundance (fmol/μg) | Median Predicted Flux (mmol/gDW/h) | Spearman ρ | p-value |
|---|---|---|---|---|---|
| ACK | Acetate kinase | 125.4 ± 15.2 | 1.85 ± 0.31 | 0.89 | < 0.001 |
| PFK | Phosphofructokinase | 88.7 ± 9.8 | 5.42 ± 0.87 | 0.92 | < 0.001 |
| MDH | Malate dehydrogenase | 64.2 ± 7.1 | 0.98 ± 0.21 | 0.45 | 0.12 |
| G6PDH | Glucose-6-P dehydrogenase | 42.5 ± 5.3 | 2.15 ± 0.44 | 0.85 | < 0.001 |
Workflow Visualization
Workflow for Proteomic-Flux Correlation Analysis
Critical Experimental Design Considerations
- Cohort Selection and Power Analysis: Ensure biological replicates (minimum n=5) to achieve sufficient statistical power for correlation analysis. Underpowered studies fail to detect significant correlations [70].
- Sample Randomization and Blinding: Randomize sample processing order during proteomic preparation and data acquisition to avoid batch effects. Implement blinding where possible to prevent analytical bias [70].
- Model Selection and Consensus Building: Be aware that different GEM reconstruction tools (CarveMe, gapseq, KBase) yield models with varying reaction content and metabolic functionality. Employ consensus approaches to generate more comprehensive and accurate metabolic networks [27].
- Validation Workflow: Adopt a sequential strategy using untargeted proteomics for discovery and hypothesis generation, followed by targeted proteomics for precise validation and quantification of key metabolic enzymes [67].
The integration of quantitative proteomic data with genome-scale metabolic modeling provides a powerful methodology for validating predicted metabolic fluxes in synthetic microbial communities. The protocols outlined herein enable researchers to move beyond purely in silico predictions toward experimentally validated models with enhanced predictive power. This correlation framework establishes a critical bridge between computational modeling and experimental measurement, ultimately accelerating the rational design of microbial consortia for therapeutic applications, drug development, and systems pharmacology. Future advancements will require continued refinement of both proteomic quantification methods and metabolic modeling algorithms to better capture the dynamic complexities of microbial community interactions.
Within the framework of comparative metabolic modeling for synthetic microbial community (SynCom) research, the transition from in silico predictions to in vivo validation represents a critical step. The functional validation of designed SynComs in gnotobiotic animal models provides an indispensable, controlled system to test hypotheses regarding community stability, host-microbe interactions, and causal mechanisms in disease. This application note details a specific case study that employs this methodology, focusing on a SynCom designed to model the inflammatory bowel disease (IBD) microbiome [4]. We outline the experimental workflow, from the community's function-based design through to its validation in gnotobiotic mice, and provide a curated toolkit of reagents and protocols to support replication of this approach.
The development and validation of the IBD SynCom followed a structured workflow, integrating computational design with in vivo experimentation. The process, summarized in the diagram below, ensures that the community selected is both functionally representative and experimentally tractable.
Function-Based Design Strategy
The IBD SynCom was constructed using a functionally directed selection strategy, prioritizing metabolic and ecological functions over purely taxonomic representation [4]. The table below summarizes the core computational steps and their objectives.
Table 1: Core Steps in the Function-Based Design of the IBD SynCom
| Step | Method/Tool | Key Objective | Output |
|---|---|---|---|
| 1. Metagenomic Analysis | MEGAHIT assembly, Prodigal, HMMscan [4] | Identify microbial functions enriched in diseased versus healthy states. | Binarized Pfam protein family vectors for metagenomes. |
| 2. Strain Selection | MiMiC2 pipeline [4] | Select isolate genomes that recapitulate the functional profile of the target ecosystem. | A candidate list of strains from a genome collection (e.g., HiBC). |
| 3. Weighting Functions | Fischer's exact test, prevalence analysis [4] | Up-weight functions that are core to the ecosystem or differentially enriched in disease. | A weighted scoring system for strain selection. |
| 4. Metabolic Modeling | GapSeq, BacArena [4] | Provide in silico evidence for cooperative strain coexistence prior to experimental validation. | Genome-scale metabolic models (GEMs) and growth simulations. |
Experimental Protocols
Protocol:In VivoValidation in Gnotobiotic Mice
This protocol details the procedure for colonizing gnotobiotic mice with the SynCom and assessing its impact on host health.
Materials and Preparation
- Gnotobiotic IL10⁻/⁻ Mice: Germ-free mice genetically susceptible to colitis [4] [54].
- SynCom Inoculum: The defined 10-member bacterial community, cultured individually and mixed in predetermined proportions [4].
- Anaerobic Workstation: For maintaining and processing anaerobic bacterial cultures.
- Diet: Standard autoclaved chow diet.
Colonization and Monitoring
- Pre-colonization: House germ-free IL10⁻/⁻ mice in flexible-film isolators to maintain sterility.
- Inoculum Preparation: Grow each SynCom member to mid-log phase in appropriate anaerobic media. Combine strains to form the final consortium.
- Oral Gavage: Administer a single dose (e.g., 200 µL) of the prepared SynCom to mice via oral gavage [54].
- Monitoring: House colonized mice in isolators and monitor for the duration of the experiment (typically 4-8 weeks). Collect fecal samples regularly to verify stable colonization via 16S rRNA sequencing or qPCR.
Phenotypic Assessment of Colitis
At the experimental endpoint, assess colitis using the following quantitative and qualitative measures:
Table 2: Key Metrics for Assessing Colitis in the Gnotobiotic Mouse Model
| Metric Category | Specific Measures | Method of Assessment |
|---|---|---|
| Clinical & Macroscopic | Body weight change, colon length, spleen weight | Calipers, weighing scale |
| Histological | Inflammatory cell infiltration, epithelial hyperplasia, crypt damage | H&E staining of colon sections, blinded histological scoring |
| Molecular | Expression of pro-inflammatory cytokines (e.g., TNF-α, IL-6, IFN-γ) | RT-qPCR on colon tissue or protein immunoassay |
In the featured case study, the 10-member IBD SynCom successfully induced colitis in the gnotobiotic IL10⁻/⁻ mice, thereby validating its functional capacity to model a disease-associated microbiome [4].
The Scientist's Toolkit
The following table compiles essential research reagents and solutions critical for the design, construction, and validation of SynComs as illustrated in the case study.
Table 3: Research Reagent Solutions for SynCom Development and Validation
| Reagent / Solution | Function / Application | Example / Source |
|---|---|---|
| Genome Collections | Source of isolate genomes for SynCom assembly. | Human Intestinal Bacterial Collection (HiBC) [4] |
| MiMiC2 Bioinformatics Pipeline | Automated, function-based selection of SynCom members from genome collections. | Custom Python scripts for Pfam vector comparison [4] |
| GapSeq | Tool for the automated reconstruction of genome-scale metabolic models (GEMs). | Used to generate metabolic models from isolate genomes [4] |
| BacArena | Toolkit for dynamic, spatially-resolved metabolic modeling of microbial communities. | Used for in silico simulation of SynCom coexistence [4] |
| Defined Microbial Media | For cultivating individual SynCom members under anaerobic conditions. | AF Medium (for OMM12 community) [71] |
| Gnotobiotic Animal Facilities | Provides a sterile environment for housing and experimenting on germ-free animals. | Flexible-film isolators for mouse studies [4] [54] |
Metabolic Modeling and Ecological Principles
The success of the described SynCom relies on computational predictions of stability, which are grounded in ecological principles and metabolic modeling. Key concepts like Metabolic Interaction Potential (MIP) and Metabolic Resource Overlap (MRO) are critical for evaluating potential coexistence in vivo [6].
Strains with a narrow spectrum of resource utilization have been shown to increase MIP and reduce MRO, thereby favoring stable metabolic interactions and coexistence within the community [6]. Furthermore, engineering SynComs with a balance of cooperative and competitive interactions, while being mindful of "cheating" behavior, helps ensure long-term resilience [2]. The strategic inclusion of keystone species that play a central role in the metabolic network further enhances structural integrity [2] [6]. Adherence to these principles during the design phase, guided by metabolic modeling, significantly increases the probability of the SynCom forming a stable community in vivo.
This application note provides a detailed protocol for assessing the predictive power of metabolic models in forecasting emergent properties in synthetic microbial communities (SynComs). It outlines standardized methods for quantifying two key emergent properties: metabolite exchange and community productivity. The protocols leverage genome-scale metabolic models (GEMs) and flux balance analysis (FBA) to simulate interactions, complemented by experimental validation workflows. Designed for researchers engaged in comparative metabolic modeling, this document facilitates the systematic evaluation of model accuracy in predicting community-level behaviors from individual strain data.
Synthetic microbial communities provide a tractable model system for uncovering the organizational principles of complex microbial ecosystems [72]. A major challenge in the field is the ability to predict emergent properties—such as stable community composition, metabolic cross-feeding, and overall productivity—that arise from multi-species interactions and are not evident from studying individual members in isolation [73]. Computational modeling, particularly constraint-based metabolic modeling, offers a powerful framework for predicting these properties in silico [27] [12].
This document presents an integrated protocol for evaluating how well different modeling approaches predict metabolite exchange and productivity. It is situated within a broader thesis on comparative metabolic modeling, aiming to provide a standardized benchmark for assessing model performance and guiding the selection of appropriate computational tools for SynCom design [27] [2].
The table below summarizes key quantitative findings from recent studies on model prediction and community assembly.
Table 1: Quantitative Data on Community Predictions and Model Performance
| Metric | Value / Finding | Context / Condition | Source |
|---|---|---|---|
| Community Stabilization | ~5 growth cycles | 10-strain SynCom in two media | [72] |
| Jaccard Similarity (Reactions) | 0.23 - 0.24 | Between GEMs from different reconstruction tools | [27] |
| Jaccard Similarity (Metabolites) | ~0.37 | Between GEMs from different reconstruction tools | [27] |
| Temporal Prediction Horizon | Up to 10 time points (2-4 months) | Graph Neural Network on WWTP data | [74] |
| Stability Assessment in Studies | ~40% (35/86 studies) | Percentage of SynCom studies evaluating stability | [2] |
Table 2: Performance Comparison of GEM Reconstruction Tools
| Tool | Reconstruction Approach | Key Characteristic | Impact on Prediction |
|---|---|---|---|
| CarveMe | Top-Down | Highest number of genes in models | [27] |
| gapseq | Bottom-Up | Largest number of reactions and metabolites; more dead-end metabolites | [27] |
| KBase | Bottom-Up | Similar reaction/metabolite sets to gapseq due to shared ModelSEED database | [27] |
| Consensus | Hybrid | Combines outputs from multiple tools; more reactions & metabolites; fewer dead-end metabolites | [27] |
Experimental Protocols
Protocol 1:In SilicoPrediction of Metabolic Exchange
Objective: To predict potential metabolite exchanges in a SynCom using Genome-Scale Metabolic Models (GEMs) and Flux Balance Analysis (FBA).
Materials:
- Genome sequences for all member strains of the SynCom.
- Computational resources (high-performance computing cluster recommended).
- GEM reconstruction tools (CarveMe, gapseq, KBase).
- Constraint-based modeling software (e.g., COBRA Toolbox).
Method:
- GEM Reconstruction: Independently reconstruct a draft GEM for each strain in the SynCom using at least two different automated tools (e.g., CarveMe and gapseq) [27].
- Model Curation: Convert all models to a consistent metabolite and reaction namespace (e.g., using MetaNetX) to enable integration [27].
- Community Model Formulation: Create a compartmentalized community model. This model combines the individual GEMs into a single stoichiometric matrix, with each species in its own compartment, linked by a shared extracellular compartment [27].
- Simulation Setup: Define a constraint-based modeling problem for the community. Set the objective function, often to maximize community biomass or the growth of a specific member.
- Flux Balance Analysis: Perform FBA and related techniques (e.g., parsimonious FBA) to simulate growth and metabolic flux distributions under defined environmental conditions [72] [12].
- Exchange Analysis: Identify metabolite exchange by analyzing the flux through transport and exchange reactions between individual models and the shared extracellular space.
Protocol 2: Experimental Validation of Emergent Properties
Objective: To empirically measure metabolite exchange and community productivity for comparison with model predictions.
Materials:
- Pure cultures of all bacterial strains in the SynCom.
- Defined minimal media (e.g., M9 or similar).
- Anaerobic chamber (for obligate anaerobes).
- Spectrophotometer or plate reader for measuring optical density (OD).
- LC-MS/MS or GC-MS for targeted metabolomics.
Method:
- Community Assembly & Passaging:
- Inoculate a defined SynCom (e.g., 10 strains from the Populus rhizosphere) into the chosen media [72].
- Serially passage the community by transferring a small inoculum (e.g., 1%) into fresh media at set time intervals or upon reaching stationary phase.
- Continue passaging for a minimum of 5 cycles to allow the community to stabilize [72].
- Productivity and Composition Monitoring:
- Track community productivity by measuring OD600 or total protein content at each passage.
- Monitor community structure by plating on selective media or via 16S rRNA amplicon sequencing to determine the relative abundance of each strain as it stabilizes.
- Metabolite Exchange Validation:
- Collect cell-free supernatant samples from the SynCom and from axenic cultures of each strain at multiple growth phases.
- Perform targeted metabolomics to identify and quantify metabolites present in the supernatant of the SynCom but absent or at lower concentrations in axenic cultures, indicating cross-fed metabolites [72] [73].
- Data Integration:
- Compare the experimentally measured emergent properties (stable composition, metabolite exchanges) with the in silico predictions from Protocol 1 to assess the predictive power of the models.
Workflow and Pathway Diagrams
SynCom Analysis Workflow
Metabolic Interaction Types
This diagram illustrates the primary metabolic interactions that can be predicted and measured.
The Scientist's Toolkit
Table 3: Essential Research Reagents and Computational Tools
| Item Name | Function / Application | Specific Example / Note |
|---|---|---|
| GEM Reconstruction Tools | Automated construction of genome-scale metabolic models from genomic data. | CarveMe (top-down), gapseq (bottom-up), KBase (bottom-up) [27] |
| Consensus Modeling | Integrates outputs from multiple reconstruction tools to reduce bias and create more comprehensive models. | Merges draft models from CarveMe, gapseq, and KBase; reduces dead-end metabolites [27] |
| Constraint-Based Modeling | Simulates metabolic flux within and between organisms in a community. | COBRA Toolbox; used for Flux Balance Analysis (FBA) [12] |
| COMMIT | A computational pipeline for gap-filling and refining community metabolic models. | Used with an iterative, abundance-based approach for model building [27] |
| Defined Microbial Strains | The foundational building blocks for constructing a SynCom with known genotypes. | e.g., 10 strains from the Populus deltoides rhizosphere [72] |
| Metaproteomics | Quantifies protein expression in a community, providing functional insights for model validation. | Used to characterize the metabolic state of a stable community [72] |
| Graph Neural Networks | A machine learning approach for predicting temporal dynamics of microbial communities. | "mc-prediction" workflow for forecasting species abundance [74] |
The Role of Agent-Based Modeling in Simulating Spatio-Temporal Community Dynamics
Application Note: Integrating ABM with Metabolic Models for SynCom Design
Theoretical Foundation and Rationale
Agent-Based Modeling (ABM) represents a paradigm shift in computational ecology by enabling researchers to simulate complex systems from the ground up, where global patterns emerge from individual interactions [75]. When applied to synthetic microbial communities (SynComs), ABM provides a powerful framework for bridging the gap between genome-scale metabolic predictions and observed spatio-temporal community dynamics. This integration is particularly valuable for addressing the persistent challenge of achieving both functional precision and ecological stability in engineered communities [2].
The core strength of ABM lies in its ability to represent autonomous, decision-making agents (individual microbial cells or populations) that interact with each other and their environment within explicitly defined spatial and temporal contexts [75]. This approach aligns perfectly with the need to model microbial interactions—including mutualism, competition, and cheating behavior—that fundamentally shape community assembly and function [2]. Recent research has demonstrated that narrow-spectrum resource-utilizing bacteria enhance community stability through increased metabolic interactions and reduced resource competition, patterns that ABM is uniquely positioned to explore mechanistically [6].
Key Integration Points with Metabolic Modeling
The synergy between ABM and metabolic modeling occurs at multiple biological scales:
- Individual-Level Metabolism: Genome-scale metabolic models (GMMs) provide the biochemical constraints that govern agent behavior, including substrate preferences, metabolic capabilities, and secretion profiles [6].
- Population-Level Interactions: ABM translates metabolic predictions into spatial interaction rules, simulating cross-feeding, competition for nutrients, and chemical warfare [2].
- Community-Level Emergence: The framework captures how localized interactions give rise to system-wide properties such as stability, resilience, and metabolic efficiency [2] [76].
Table 1: Quantitative Metrics for ABM of SynComs
| Metric Category | Specific Parameters | Theoretical Basis |
|---|---|---|
| Spatial Structure | Spatial aggregation index, Local hotspot density | SVGbit computational pipeline for spatial patterns [77] |
| Metabolic Interactions | Metabolic Interaction Potential (MIP), Metabolic Resource Overlap (MRO) | Genome-scale metabolic modeling [6] |
| Community Stability | Resistance, Resilience, Robustness | Ecological stability theory [2] |
| Agent Properties | Resource utilization width, Phylogenetic distance | Phenotype microarray data [6] |
Protocol: ABM Development Workflow for Microbial Community Dynamics
Agent and Environment Specification
This protocol outlines a standardized workflow for developing ABM simulations of synthetic microbial communities, with particular emphasis on integration with metabolic modeling data.
Step 1: Agent Definition and Parameterization
- Isolate Core Functional Traits: Define agent properties based on experimentally determined strain characteristics, including: (1) resource utilization profiles from phenotype microarrays [6]; (2) metabolic interaction potential (MIP) scores from GMM simulations [6]; and (3) functional capabilities (e.g., nitrogen fixation, phosphate solubilization) [6].
- Implement Behavioral Rules: Program agent decision-making based on ecological principles, including:
Step 2: Environment Configuration
- Define Spatial Dimensions: Establish a simulated environment reflecting target habitats (e.g., rhizosphere soil particles, biofilm surfaces) with appropriate scale and boundary conditions [75].
- Parameterize Resource Gradients: Implement dynamic nutrient distributions based on target environments, including carbon sources identified through phenotype arrays [6].
Simulation Implementation and Validation
Step 3: Interaction Rule Implementation
- Encode metabolic constraints using vector-agent principles where agents modify their geometric attributes (position, metabolic state) based on local conditions [75].
- Implement dynamic interaction networks that evolve based on:
- Local nutrient concentrations
- Neighbor agent states and types
- Temporal factors (growth phase, accumulation of waste products)
Step 4: Model Validation and Calibration
- Pattern-Oriented Modeling: Compare emergent simulation patterns with experimental data using multiple quantitative metrics [75]:
- Spatial validation: Compare simulated spatial aggregation with empirical spatial transcriptomics data (e.g., from SVGbit analysis) [77]
- Temporal validation: Match population dynamics with observed growth curves and community succession patterns
- Functional validation: Verify that emergent metabolic interactions align with GMM predictions [6]
Step 5: Scenario Testing and Analysis
- Perturbation experiments: Simulate environmental stressors (antibiotic exposure, nutrient limitation) to assess community stability [2].
- Keystone species manipulation: Test the impact of removing specific narrow-spectrum resource-utilizing strains predicted to stabilize communities [6].
- Long-term adaptation: Incorporate evolutionary dynamics to model trait evolution under sustained selection pressures [2].
Table 2: Research Reagent Solutions for ABM-SynCom Integration
| Reagent/Resource | Function in Workflow | Implementation Example |
|---|---|---|
| Phenotype Microarrays | Quantifies resource utilization spectra | Determines agent metabolic capabilities and niche width [6] |
| Genome-Scale Metabolic Models (GMMs) | Predicts metabolic interactions | Parameterizes cross-feeding rules and competition dynamics [6] |
| Spatial Transcriptomics Data | Provides empirical spatial patterns | Validation benchmark for simulated spatial organization [77] |
| Convolutional Non-negative Matrix Factorization (CNMF) | Identifies spatiotemporal motifs | Analyzes simulated activity patterns for recurrent dynamics [78] |
| Vector-Agent Modeling Framework | Represents geometric autonomy | Enables realistic spatial movement and interaction [75] |
Protocol: Advanced Analysis of Emergent Spatio-Temporal Patterns
Dimensionality Reduction and Motif Analysis
Objective: Identify recurrent spatio-temporal patterns in simulated community dynamics that correspond to core ecological processes.
Procedure:
- Simulation Output Processing: Export agent states and environmental conditions at regular intervals (e.g., every simulated hour) to create a spatio-temporal data cube.
- Pattern Mining: Apply convolutional non-negative matrix factorization (CNMF) to identify recurrent activity motifs—characteristic patterns of community-wide metabolic activation or spatial organization [78].
- Dimensionality Assessment: Quantify the complexity of community dynamics by determining the number of motifs required to explain >90% of variance in system behavior—typically ~14 core patterns in stable systems [78].
Interpretation Guidelines:
- Low-dimensional dynamics (few motifs explaining high variance) indicate stable, predictable community states
- Motif engagement shifts in response to perturbations signal community resilience or fragility
- Context-specific motifs that activate under particular conditions represent specialized functional responses
Stability and Resilience Quantification
Objective: Apply rigorous metrics to assess community stability properties emerging from ABM simulations.
Procedure:
- Baseline Establishment: Run simulations under constant conditions to establish stable reference states for each SynCom configuration.
- Perturbation Application: Introduce disturbances including:
- Pulse perturbations: Temporary antibiotic exposure or nutrient pulses
- Press perturbations: Sustained environmental changes (pH shifts, temperature changes)
- Stability Metric Calculation:
- Resistance: Quantify as (1 - [maximum deviation from baseline]/[perturbation magnitude])
- Resilience: Calculate as recovery rate to baseline following perturbation
- Robustness: Measure as maintenance of specific functions under disturbance [2]
Integration with Experimental Validation:
- Compare ABM-predicted stability metrics with empirical measurements from SynCom experiments in target environments [6]
- Use tipping point analysis to identify critical thresholds in environmental parameters that trigger community collapse [76]
- Validate predicted keystone species roles through targeted removal experiments [6]
Table 3: Stability Optimization Strategies for SynCom Design
| Design Strategy | Mechanism | ABM Implementation |
|---|---|---|
| Narrow-Spectrum Resource Utilization | Reduces metabolic resource overlap (MRO) | Agent metabolic specialization rules [6] |
| Interaction Network Balancing | Dynamic equilibrium of cooperative and competitive relationships | Weighted interaction probabilities [2] |
| Keystone Species Governance | Structural integrity through influential species | Differential agent influence parameters [2] |
| Modular Metabolic Stratification | Efficient resource partitioning | Spatial zoning of metabolic functions [2] |
| Evolution-Guided Selection | Overcoming functional-stability trade-offs | Multi-generational trait inheritance [2] |
Concluding Remarks
The integration of Agent-Based Modeling with metabolic theory represents a transformative approach to designing synthetic microbial communities with predictable dynamics. By bridging genomic capabilities with emergent spatio-temporal patterns, this framework addresses the fundamental challenge of achieving both functional precision and ecological stability in engineered systems [2] [6].
The protocols outlined here provide researchers with practical methodologies for implementing ABM that is firmly grounded in experimental data and metabolic constraints. This approach enables the exploration of design principles such as the strategic inclusion of narrow-spectrum resource-utilizing strains to enhance stability through increased metabolic interaction potential [6]. Furthermore, the emphasis on spatio-temporal motif analysis creates opportunities for identifying universal patterns in microbial community organization across different habitats and functions [78] [77].
As synthetic biology continues to advance toward more complex multicellular systems, ABM will play an increasingly critical role in predicting how engineered functions manifest in spatially structured, dynamic environments. The integration of machine learning with the framework described here promises to further accelerate the design-build-test-learn cycle, ultimately enabling the programming of microbial communities as ecotechnologies for addressing global sustainability challenges [2].
Conclusion
Comparative metabolic modeling has matured into a powerful, indispensable framework for transitioning from descriptive ecology to predictive engineering of synthetic microbial communities. By integrating foundational ecological principles with advanced computational methods like consensus GEMs and machine learning, researchers can now navigate the complexities of higher-order interactions and design stable, functionally robust SynComs. The convergence of top-down and bottom-up design strategies, validated through rigorous in silico and in vivo models, paves the way for transformative biomedical applications. Future efforts must focus on standardizing model reconstruction, exploiting microbial dark matter with AI, and developing digital twins to accurately simulate host-microbe dynamics. This progression will ultimately enable the reliable deployment of bespoke microbial consortia for targeted therapeutic interventions, personalized microbiome-based drugs, and the sustainable engineering of complex biological systems.
References
- 1. Synthetic microbial communities: Bridging research and ... [https://link.springer.com/article/10.1007/s11104-025-07937-y]
- 2. Ecological design of high-performance synthetic microbial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12373479/]
- 3. The Role of Synthetic Microbial Communities (SynCom) in ... [https://www.frontiersin.org/journals/agronomy/articles/10.3389/fagro.2022.896307/full]
- 4. Function-based selection of synthetic communities enables ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12507024/]
- 5. Data-driven synthetic microbes for sustainable future [https://www.nature.com/articles/s41540-025-00556-4]
- 6. Narrow-spectrum resource-utilizing bacteria drive the ... [https://www.nature.com/articles/s41467-025-61432-7]
- 7. Keystone Species - National Geographic Education [https://education.nationalgeographic.org/resource/keystone-species/]
- 8. Metabolic Interdependence Network → Area [https://lifestyle.sustainability-directory.com/area/metabolic-interdependence-network/]
- 9. Metabolic interdependencies in thermophilic communities are ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11408653/]
- 10. Contribution of genome‐scale metabolic modelling to ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9324083/]
- 11. Using metabolic networks to resolve ecological properties ... [https://www.sciencedirect.com/science/article/pii/S245231001730197X]
- 12. Metabolic modeling of host-microbe interactions [https://www.sciencedirect.com/science/article/pii/S2001037025004076]
- 13. analysis of Comparative of metabolic ... models microbial [https://pmc.ncbi.nlm.nih.gov/articles/PMC11116368/]
- 14. Bottom-Up Approaches to Synthetic Cooperation in Microbial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6462982/]
- 15. How to Count Our Microbes? The Effect of Different ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7426659/]
- 16. Drug-microbiota interactions: an emerging priority for ... [https://www.nature.com/articles/s41392-023-01619-w]
- 17. Principles and Terminology for Host–Microbiome–Drug ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10167991/]
- 18. The Use of Defined Microbial Communities To Model Host ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6684003/]
- 19. Fast quantification of gut bacterial species in cocultures ... [https://www.nature.com/articles/s43705-022-00123-6]
- 20. Deciphering and designing microbial communities by ... [https://www.sciencedirect.com/science/article/pii/S2001037024001387]
- 21. Synthetic microbial community maintains the functional ... [https://www.sciencedirect.com/science/article/abs/pii/S0043135425001848]
- 22. Interactions between native soil microbiome and a ... [https://pubmed.ncbi.nlm.nih.gov/40852997/]
- 23. Metabolic Modeling of Microbial Community Interactions for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6225453/]
- 24. Genome-Scale Metabolic Modeling Enables In-Depth ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8778254/]
- 25. A metabolic modeling platform for the computation of ... [https://www.nature.com/articles/s41596-021-00593-3]
- 26. A protocol for generating a high-quality genome-scale ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3125167/]
- 27. Comparative analysis of metabolic models of microbial ... [https://www.nature.com/articles/s41540-024-00384-y]
- 28. Higher-order microbial interactions revealed by ... [https://pubmed.ncbi.nlm.nih.gov/40937471/]
- 29. Genome scale metabolic network modelling for metabolic ... [https://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.1011381]
- 30. Recent advances on constraint-based models by ... [https://www.sciencedirect.com/science/article/pii/S095816691930117X]
- 31. Simulated Design–Build–Test–Learn Cycles for Consistent ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10510747/]
- 32. DBTL bioengineering cycle for part characterization and ... [https://www.sciencedirect.com/science/article/pii/S2405896324017488]
- 33. The knowledge driven DBTL cycle provides mechanistic ... [https://microbialcellfactories.biomedcentral.com/articles/10.1186/s12934-025-02729-6]
- 34. iNAP 2.0: Harnessing metabolic complementarity in microbial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11487609/]
- 35. Multifaceted Applications of Synthetic Microbial Communities [https://www.mdpi.com/2036-7481/15/3/113]
- 36. Metabolic modelling approaches for describing and ... [https://www.sciencedirect.com/science/article/pii/S2001037020305286]
- 37. OptCom: A Multi-Level Optimization Framework for the ... [https://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.1002363]
- 38. Dynamic flux balance analysis for synthetic microbial ... [https://pubmed.ncbi.nlm.nih.gov/25257022/]
- 39. Designing a synthetic microbial community through genome ... [https://environmentalmicrobiome.biomedcentral.com/articles/10.1186/s40793-023-00536-3]
- 40. Revealing the dynamics and mechanisms of bacterial ... [https://www.sciencedirect.com/science/article/pii/S1096717624000302]
- 41. Accurate profiling of microbial communities for shotgun ... [https://microbiomejournal.biomedcentral.com/articles/10.1186/s40168-025-02249-w]
- 42. Network-based multi-omics integrative analysis methods in ... [https://biodatamining.biomedcentral.com/articles/10.1186/s13040-025-00442-z]
- 43. Integration of Omics Data Sources to Inform Mechanistic ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7158209/]
- 44. Advancing drug development with “Fit-for-Purpose” modeling ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12436579/]
- 45. An Efficient, Low-cost and Scalable Method for ... [https://bio-protocol.org/exchange/protocoldetail?id=2405576&type=1]
- 46. gapseq: informed prediction of bacterial metabolic pathways ... [https://genomebiology.biomedcentral.com/articles/10.1186/s13059-021-02295-1]
- 47. Genome-scale metabolic reconstruction of 7302 human ... [https://www.nature.com/articles/s41587-022-01628-0]
- 48. Bactabolize: A tool for high-throughput generation of ... [https://elifesciences.org/reviewed-preprints/87406]
- 49. Bactabolize is a tool for high-throughput generation of ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10564454/]
- 50. GEMsembler: consensus model assembly and structural ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12542698/]
- 51. Towards synthetic ecology: strategies for the optimization ... [https://www.frontiersin.org/journals/synthetic-biology/articles/10.3389/fsybi.2025.1532846/full]
- 52. COMMIT: Consideration of metabolite leakage and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8942231/]
- 53. Teasing out missing reactions in genome-scale metabolic ... [https://www.nature.com/articles/s41467-023-38110-7]
- 54. Design and application of synthetic human gut microbial ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12622336/]
- 55. Synthetic microbial communities for studying and ... [https://www.sciencedirect.com/science/article/pii/S136952742500058X]
- 56. A systematic discussion and comparison of the construction methods... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11253132/]
- 57. Interspecific competition prevents the proliferation of social ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10939377/]
- 58. Bottom-Up Approaches to Synthetic Cooperation in ... [https://www.mdpi.com/2075-1729/9/1/22]
- 59. Full factorial construction of synthetic microbial communities [https://elifesciences.org/reviewed-preprints/101906]
- 60. Current status and applications of genome-scale metabolic ... [https://genomebiology.biomedcentral.com/articles/10.1186/s13059-019-1730-3]
- 61. Automated design of synthetic microbial communities [https://www.nature.com/articles/s41467-020-20756-2]
- 62. Synthetic Microbial Ecology: Engineering Habitats for ... [https://www.frontiersin.org/journals/microbiology/articles/10.3389/fmicb.2017.01125/full]
- 63. Comparison of metabolic states using genome-scale ... [https://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.1009522]
- 64. Visualizing metabolic network dynamics through time-series ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7119163/]
- 65. MetaboTools: A Comprehensive Toolbox for Analysis of ... [https://www.frontiersin.org/journals/physiology/articles/10.3389/fphys.2016.00327/full]
- 66. Benchmarking Procedures for High-Throughput Context ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC4722106/]
- 67. Application of proteomic data in the translation of in vitro ... [https://www.sciencedirect.com/science/article/abs/pii/S174067492100010X]
- 68. Flux sampling in genome-scale metabolic modeling of ... [https://bmcbioinformatics.biomedcentral.com/articles/10.1186/s12859-024-05655-3]
- 69. Finding a vocation for validation: taking proteomics beyond ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC9906375/]
- 70. Tutorial: best practices and considerations for mass ... [https://www.nature.com/articles/s41596-021-00566-6]
- 71. In vitro interaction network of a synthetic gut bacterial ... [https://www.nature.com/articles/s41396-021-01153-z]
- 72. Formation, characterization and modeling of emergent ... [https://www.sciencedirect.com/science/article/pii/S2001037021001069]
- 73. To Study Competition and Cross-Feeding, Scientists Build ... [https://www.energy.gov/science/ber/articles/study-competition-and-cross-feeding-scientists-build-synthetic-microbiomes]
- 74. Predicting microbial community structure and temporal ... [https://www.nature.com/articles/s41467-025-64175-7]
- 75. A vector-agent approach to (spatiotemporal) movement ... [https://www.nature.com/articles/s41598-022-22056-9]
- 76. An agent-based model of the extinction of experience [https://www.sciencedirect.com/science/article/pii/S0272494425001124]
- 77. The spatiotemporal dynamics of spatially variable genes in ... [https://www.nature.com/articles/s41420-023-01569-w]
- 78. Low-Dimensional Spatio-Temporal Dynamics Underlie Cortex ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC7375907/]